HOOFDSTUK 2: DE CEL: MORFOLOGISCHE EN FUNCTIONELE EENHEID VAN LEVEN
2.1 Observeerbaarheid, organisatie en eigenschappen van het plasmalemma
Alle cellen zijn minimaal omgeven met een plasmalemma
Over het plasmalemma gebeurt communicatie en uitwisseling met extracellulair milieu
Plantencellen hebben celwand
Fosfolipiden: Hydrofiel hoofd en hydrofobe staart
dubbele fosfolipidenlaag, staarten nr elkaar toe
Naast fosfolipiden ook cholesterol
Specifieke membraaneiwitten verankerd in membraan met transmembranair domein, ook
intra/extracellulaire domeinen
Functie membraaneiwitten: receptor, signaaltransductie, kanaal, pomp
Sommige geen vaste locatie, andere verankerd via cytoskeletelementen (vloeistofmoziekmodel)
Zelfsluitbaar
Lipofiele basis flexiebel, rekbaar, vormwijzigingen mogelijk
Afsplitsbaarheid (bij cytokinese of celdeling)
Versmeltbaarheid en afsplitsbaarheid maken endo- en exocytose mogelijk
Niet permeabel vr ionen, maar transport geregeld door transmembranaire eiwitten die
fungeren als kanaal of pomp
H2O verplaatst zich door thermische agitatie
Selectieve permeabiliteit
Diffusie: passieve verplaatsing van opgeloste stoffen volgens hun concentratiegradiënt
Osmose: passieve verplaatsing van oplosmiddel van hypotonsich nr hypertonisch
comparti- ment tot de situatie isotonisch geworden is
Osmotische druk: de op een oplossing uit te oefenen hydrostatische druk die wateraanzuiging vanuit een naburig maar door semi-permeabel membraan gescheiden hypotonisch compartiment neutraliseert
Elektrische dimensie van het plasmalemma
Cellen continu onderhevig aan diffusie en osmose (tenzij in isotonisch milieu)
Intracellulair hoge K+ concentratie, extracellulair hoge Na+ concentratie
Theoretisch: K+ ionen lekker nr buiten en Na+ ionen diffunderen naar binnen tot evenwicht
In realiteit: ionenpompen in het plasmalemma die evenwichtssituatie proberen te herstellen
Bij verplaatsing van een ion wordt deze verplaatsing tegengewerkt dr potentiaalverschil
Bij evenwicht blijft er evenwichtsmembraanpotentiaal zodat membraan gepolariseerd is
(binnen negatief, buiten positief)
Membraan alleen permeabel vr X
Verplaatsing tot evenwicht
Ontstaan elektrische gradiënt, potentiaalverschil
X+
X+
+
X
X+YX+Y-
Zenuwcel
Na+
K+-kanaal open, K+ lekt nr buiten
K
+
Plots neemt permeabiliteit vr Na+ toe, dus Na+ nr binnen, minder
negatief = depolarisatie
-70mV
Nernstvergelijking
Em = RT ln pK+ [K+]o + pNa+ [Na+]o + pCl-[Cl-]i
ZF
pK+ [K+]i + pNa+ [Na+]i + pCl-[Cl-]o
Em = 58 log10 1[20] + 0,01 [440] + 0,01 [100]
1[400] + 0,01 [50] + 0,01[560]
= 58 log10 [25] = -70mV (binnen negatief tov buiten)
[406]
Opm.: Em = Em K+ permeabiliteit voor K+ is 100 keer groter, dus andere concentraties
verwaarloosbaar bij evenwicht
Logaritmische functie!
Uitwendig 10 K+, inwendig 100 K+ (depolarisatie):
Em = 58 log 10 = 58 (log10 – log 100) = -58 mV
100
Als inwendige en uitwendige concentratie gelijk zijn, E m = 0
Belangrijkste functie van het plasmalemma is communicatie
Op membraan receptor waaraan ligand kan binden, via intermediaire eiwitten wordt de informatie
dan getransporteerd
Onderlinge communicatie tss cellen via hormonen
2.2 Intracellulaire organisatie van eukaryote cellen
RIBOSOMEN :
Vrij in cytoplasma of verankerd aan membranen van het RER
Associatie van ribosomale RNA’s en ribosomale eiwitten
Kleine en grote subeenheid
Snoer van ribosomen = polyribosoom
Eiwitsynthese gebeurt hier, mRNA streng tss grote en kleine subeenheid
ENDOPLASMATISCH RETICULUM
Vormt systeem van kanalen, holten en cysternae
SER: steroïd synthetiserende cellen
Ca++- stockage in spiercellen (spiercontractie)
RER: ER met ribosomen
Synthese van eiwitten voor transport naar het plasmalemma voor inbouw en secretie
Verbinding naar Golgi-apparaat waar processing en condensering gebeurt, gevolgd dr in
sluiting in vesikels
GOLGI-APPARAAT
Aan de zijkant afsnoerende blaasjes
Eiwitten komen van RER, naar binnen langs CIS-Golgi
processing (plooiing door S-bruggen tss cysteïne’s, aanhechting suikers of lipiden)
vesiculair transport naar midden-Golgi
trans-golgi: eiwitten condenseren, verpakking in vesikel
naar membraan voor inbouw of secretie
MITOCHONDRIA
Dubbel membraan (endosymbiose-hypothese), binnenste geplooid in cristae
Energiewinning door oxidatieve fosforylatie
VACUOLEN
Groot bij plantencellen
Bij dierlijke cellen geen (uitzondering: vetcellen, sommige witte bloedcellen)
Als er veel kleine vacuolen te zien zijn, sterft de cel (necrosis, valt uit elkaar)
LYSOMSOMEN EN PEROXISOMEN
Lysosomen bevatten lytische enzymen (hydrolasen) die in fagocyterende cellen helpen bij
intracellulaire vertering en spelen een rol bij autofagie (opruimen versleten organellen)
Peroxisomen bevatten vrije zuurstofradicalen om een ge-endocyteerde indringer te doden
CENTROSOOM
2 centriolen loodrecht op elkaar
Centriolen: MTOC (microtubuli-organiserend centrum)
Microtubuli: opgebouwd uit α en β tubuline, vormen eerst heterodimeren, dan multimeren
ontstaan “draad” = protofilament
13 protofilamenten zijdelingse associatie microtubulus (20nm)
Microtubuli is basis voor spoelfiguur
Centriool: 9 tripletten van microtubuli
Centriolen vormen ook de microtubuli van het cytoskelet
CYTOSKELET
Vezelachtige eiwitten: - microtubuli (20nm)
- intermediaire filamenten (10nm)
- microfilamenten (5nm)
Dynamisch-structurerend karakter, geleiding, helpen bij verplaatsing dr langer/korter te worden
KERN
Omgeven dr dubbele fosfolipidendubbellaag, die continu is met membranen van ER
Kernporiën
Bevat genetische informatie: DNA
- in interfase: chromatine
- bij cel in deling: chromosomen (meer gecondenseerd en gespiraliseerd)
- solenoïd model
3 niveau’s van spiralisatie:
1. DNA-helix: 2 antiparallelle complementaire strengen
2. DNA-helix ± 2 keer gedraaid rond eiwitblok (octameren) bestaande uit histonen (4 paren)
4 histonenparen vormen een nucleosoom
De helix rond het nucleosoom vormt een chromatosoom
3. Ongeveer 6 chromatosomen rond histon 1 gespiraliseerd
In elkaar gedraaid: chromosomen: 2 chromatiden samengehouden dr centromeer
Heterochromatine: hogere densiteit, kleurt gemakkelijk met DNA kleurstoffen
Euchromatine: lagere densiteit
NUCLEOLI
Hier gebeurt RNA synthese kleuren met RNA kleurstoffen
2.3 Unicellulaire levensvormen: de cel als functionele eenheid van leven
Protozoa = 1-cellige (amoebe, pantoffeldiertje)
Geen celwand, wel versterking door suikers (glycocalyx)
Binnen korrelig cytoplasma: - hyaliene ectoplasma (geen insluitsels)
- endoplasma
Kern kan ontdubbeld zijn in micro- en macronucleus (bij pantoffeldiertje wel, amoebe niet)
Pulserende vacuolen om osmotisch naar binnen komend water weg te pompen
Indeling op voortbeweging
Pseudopodia (amoebe)
Endoplasma bestaat uit - plasmagel (buitenkant, korrelige insluitsels, beweegt niet, vast)
- plasmasol (binnenkant, beweegt wel, vloeibaar)
Plasmasol wordt continu omgevormd tot plasmagel en omgekeerd (aan voorzijde)
Undulipodia: flagella (oogdiertje)
cilia (pantoffeldiertje)
Undulipodia worden ondersteund door microtubuli: 9 dupletten en 1 centraal paar
Dyenine zijarmen zorgen ervoor dat de dupletten t.o.v. elkaar bewegen
Pass. voortb. (parasieten): plasmodium in Anophelesmug malaria (rbc barsten koorts)
Bij de mens: pseudopodia: bepaalde witte bloedcellen
flagella: spermatozoa
cilia: eileider en trachea
passieve voortbeweging: bloedcellen doorheen arteries en venen
Voeding
Heterotrofie = endocytose
Opname vast voedsel: fagocytose, vloeibaar: pinocytose
Pseudoporiën omvloeien voedsel en versmelten erachter ter vorming van een voedselvacuole of
voedsel botst tg de cel, membraan invagineert en vloeit samen achter voedseldeeltje
Voedsel is niet onmiddellijk bruikbaar vertering nodig
Voedselvacuole versmelt met lysosomen ter vorming van verteringsvacuole
Aanzuring lumen dr protonenpompen enzymen geactiveerd vertering
Bouwstenen diffunderen nr cytoplasma toe, ‘afval’ versmelt met membraan exocytose
Evolutie:
Gespecialiseerd
monden
voedselverzamelapparaat,
extracellulaire
vertering,
gecompartimentaliseerd spijsverteringskanaal
Intracellulaire vertering vr opruim van versleten organellen of eliminatie dr autofagie
Polysacchariden
Glycogeen (dierlijke reserve) en zetmeel (reserve bij planten) worden verteerd door
carbohydrasen om lichaamseigen suikers te maken
Zetmeel en glycogeen zijn polymeren van glucose
Cellulose is voor de meeste heterotrofen onverteerbaar
Eiwitten
Bestaan uit AZ-ketens
Verteerd door proteasen tot AZ n en peptiden
Lipiden
Polymeren van VZ
Vertering dr lipasen tot VZn en glycerol
Nucleïnezuren
DNA, RNA
Polymeren van nucleotiden: ribosesuiker + base + fosfaat
DNA: dubbelstrengig door basepaarvorming
Metabolisme
Anabolisme (opbouwend) & katabolisme (afbrekend)
Vertering hoort niet bij het katabolisme, komt niet in cellulair milieu
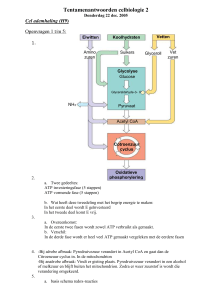
Cellulaire ademhaling
Energie uit glucosebindingen omzetten naar bruikbare energie (ATP)
Glucose C6H12O6 670 kcal
Energie is niet onmiddellijk bruikbaar, ATP wel
ATP: nucleotide met purine adenine als base, centrale ribosesuiker en een staart van 3
energierijke fosfaatbindingen
Fosfaatgroep getransfereerd nr ander molecule deze hoger E-niveau bepaalde reacties
kunnen opgaan = FOSFORYLATIE
ATP ADP + Pi + 8kcal
ADP AMP + Pi + 8kcal
Glucose wordt gefosforyleerd en dan opgesplitst in 2 x glyceraldehyde
Breken van bindingen stelt energie vrij
Energie wordt eerst gebruikt voor het reduceren van intermediaire energiedragers (niet meteen
voor aanhechting fosfaatgroep)
Intermediaire energiedragers
NAD+ + 2 e- +H2 NADH + H+
FAD + 2 e- +H2 FADH2
NADP+ + 2 e- +H2 NADPH + H+
Oxidatie van deze dragers stelt de energie weer vrij
O2 is finale e--acceptor
O2 is noodzakelijk
H2O is het eindproduct
I: GLYCOLYSE (anaëroob)
Gebeurt in het cytoplasma
Glucose wordt afgebroken tot 2 keer pyrodruivenzuur
2 glyceraldehyde moleculen als tussenproduct
~ ~
2 ATP
PGAL
2ATP
2ATP
2 NADH
C3
2ATP: ± 2% rendement
2NADH
C3
Glucose opsplitsen kan pas na fosforylering en intramoleculaire reorganisatie tot fructose 1,6difosfaat (= investering 2 ATP)
Energetische verdienste van glycolyse: geen O2 nodig
pyruvaatsubstraten waaruit E gehaald wordt via aërobe
ademhaling
II: OXIDATIE VAN PYRODRUIVENZUUR TOT ACETYL-COENZYME A
In binnencompartiment van mitochondria
Pyrodruivenzuur wordt afgebroken tot CO2 en azijnzuur (CH3COOH) dat gebonden wordt aan
coenzym A ter vorming van acetyl-coenzyme A
E wordt opgestapeld onder de vorm van 2 gereduceerde NAD moleculen
2 NADH
CO2 + CH3COOH
C2-S-CoA
III: Krebscyclus
2 acetyl-CoA moleculen worden aan oxaalazijnzuur (4C) gebonden vorming citroenzuur (6C)
wordt geoxideerd met telkens vrijstelling van CO 2
4C weer omgevormd tot oxaalazijnzuur, cyclus herbegint
E opgestapeld in 6 NADH en 2 FADH2, 2 ATP gevormd
Oxaalazijnzuur C4 + C2-S-CoA
C6
CO2
2 ATP
6 NADH
2 FADH2
C5
CO2
IV: ADEMHALINGSELECTRONENTRANSPORTKETEN
Tot nu: 10 NAD’s en 2 FAD’s :E vrijgesteld door oxidatie: O2 + 2 NADH 2 H2O + 2 NAD + E
NADH geeft E onrechtstreeks aan O2: e—transportketen met Fe-bevattende redoxmoleculen
Vrijgekomen E wordt gebruikt voor de aanmaak van ATP
Componenten van de ademhalingsketen zitten in geplooide binnenmembraan vd mitochondria
NADH en FADH2 worden geoxideerd dr e--overdracht aan 1ste pigment (flavoproteïne)
e- wordt doorgegeven met overeenkomstige redoxpotentiaal tot bij O 2 met de vorming van H2O
Bij doorlopen vd keten wordt een deel vd E overgedragen op volgend pigment en een deel
gebruikt om protonen vanuit het lumen nr de intermembranaire ruimte te pompen
Over binnenste membraan ontstaat een concentratiegradiënt en een elektrische gradiënt
NADH
e-
Verder in keten steeds hogere redoxpotentiaal
O2: finale e--accpetor
Elektrische gradiënt: negatief in matrix
pos. in intermembr. ruimte
eH+
H
ADP+Pi ATP
H
+
+
e-
H
+
e-
H+ H+
F1-complex
H+
6 O2
12 H2O
V: CHEMI-OSMOTISCHE SYNTHESE VAN ATP
Elektrische gradiënt en concentratiegradiënt H+ dr F1-complex elektrische stroom gebruikt
om ATP te maken: 32 ATP
TOTAAL: 36 ATP 34% rendement
Fotosynthese
chlorophyl
6 CO2 + 12H2O
C6H12O6 + 6 O2 + 6 H2O
Ombouw van
-energie naar stockeerbare energie (glucose) dr Groen-purperbacteria,
cyanobacteria, algae, wieren en groene planten
Organismen moeten beschikken chl-a als fotogevoelige pigmenten, zij kunnen door
elementen omzetten nr organische moleculen (autotroof vr C)
Cyanobacteria ook autotroof vr N
-energie C-
Chloroplasten: dubbel membraan (endosymbiose hypothese)
geplooid binnenmembraan, vesikels worden afgesnoerd
vesikels (thylakoïden) zijn gestapeld: grana-thylakoïden
in de thylakoïden: chlorophyl
O2 als bijproduct vrijgesteld in de atmosfeer
Planten: primaire voedselbron vr herbivore heterotrofe organismen
Samen met cyanobacteria en wieren basis vd atmosferische O 2-bron
Fotosynthese bestaat uit lichtreacties en donkerreacties. Tijdens de lichtreacties wordt de
energie gebruikt vr het genereren van ATP-moleculen. De energie wordt opgeslagen in
gereduceerde intermediairen. Tijdens de donkerreacties wordt de energie aangewend vr inbouw
van C-moleculen in organische moleculen. Dit is de snelheidsbepalende (vertragende) stap.
Lichtreacties
Twee fotosystemen in de membranen vd grana-thylakoïden
Fotosysteem I is het oermechanisme, II is evolutief gerealiseerde optimalisatie
FOTOSYSTEEM I: CYCLISCH EN A-CYCLISCH
In de wand vd grana-thylakoïden zitten lichtgevoelige pigmenten. Bij inval van een foton gaan ze
E-inhoud kanaliseren nr chl-a, P700.
P 700 gaat over nr aangeslagen toestand en gaat e- overdragen aan acceptormolecule in
membraan. Dit wordt gereduceerd en laat het e- de e--transportketen doorlopen. Het e- valt
terug op P 700, dat hierdoor hersteld wordt. De cyclus herbegint.
Tijdens het cyclisch verloop worden er protonen vanuit de matrix nr het lumen gepompt. Zo
wordt er een gradiënt gevormd. De protonen gaan dr F 1-complex waardr volgende reactie opgaat:
ADP + Pi ATP
2 aangeslagen P 700 deeltjes kunnen e- ook overdragen aan andere intermediaire redoxmoleculen;
op NADP ter vorming van NADPH + H+. Dit is het a-cyclisch verloop van fotosysteem I. P 700
blijft nu geoxideerd achter, kan niet meer deelnemen aan E-omzetting. belang fotosysteem II
FOTOSYSTEEM II: H2O ALS FINALE E- DONOR MET VRIJSTELLING VAN O2
Inval foton zorgt dat P680 aangeslagen wordt. Dit draagt e- over aan een acceptormolecule
vroeger in de e--transportketen( dus langere keten). Finaal komt het e - wel bij P 700 dat
gereduceerd wordt en bij inval van een foton weer aangeslagen kan worden.
P 680 heeft een dermate positieve redoxpotentiaal dat het e - kan wegnemen bij H2O met de
vorming van O2!!! Dus O2 afkomstig van H2O!!!
Combinatie van I en II met als onmiddellijk resultaat E opgeslagen in NADPH + H + geeft ook
verhoogde protonengradiënt waardr meer ATP geproduceerd kan worden.
ADP + Pi ATP
H+ H+
H+
NADPH + H+ NADP
Matrix chloroplast
2e-
Membr
grana
thylakoïd
-
2e
-
2e
2e
-
2e
2eChla*
Chla+ Chla
H2O
O2+2H+ P680
Foton
H+
H+
H+
Lumen
Grana
thylakoïd
fotosysteem II
2eChla*
-
H+
2e-
H+
2e-
H+
2e-
2e2e-
ATP
synthese
Chla+Chla
P 700
+
H
Foton
fotosysteem I
fotosysteem I a-cyclisch
cyclisch
Donkerreacties
Koolstof assimilatie of Calvin-Benson cyclus
C-atoom van CO2 gebonden aan ribulose bifosfaat (5C). Zo wordt een onstabiel 6C molecule
gevormd dat opgesplitst wordt in 2 3C moleculen. Deze 3C moleculen worden via aanwending van
ATP en NADPH + H+ (afkomstig vd lichtreacties) omgevormd tot PGAL (fosfoglyceraldehyde). Dit
gebeurt 3 keer 6 PGAL. 5 worden er mits investering van ATP omgevormd tot 3 ribulose
bifosfaat die opnieuw binnentreden in de cyclus. Het niet gerecycleerde PGAL wordt gebruikt in
suiker-, VZ- en AZ-metabolisme.
Vr 1 glc molecule (2PGAL) moeten de C-atomen van 6 CO2 moleculen geassimileerd worden.
Opm.: E-metabolisme lijkt onlogisch: tijdens lichtreacties ATP maken die tijdens
donkerreacties gebruikt wordt vr de synthese van glucose.
Tijdens de cellulaire ademhaling E spenderen om de in glucose opgestapelde E om te bouwen in
ATP. MAAR zelfde E opslaan onder ATP is onmogelijk, glucose is dense stockeerbare energiebron
en de assimilatiereacties zijn ook nodig vr essentiële metabolisme van andere biomoleculen.
Planten stockeren E onder de vorm van zetmeel, dieren onder de vorm van glycogeen, beide
specifieke polymeren van glucose
Anaërobe omstandigheden: alleen glycolyse opstapeling pyruvaat evenwichtsreacties v
glycolyse dreigen stil te vallen
Bacteriën, gistcellen en spiercellen hebben probleem omzeild dr eindproduct vd glycolyse
enzymatisch om te zetten (zonder O2 nodig te hebben) = verklaring proces van
alcoholfermentatie bij gisten en melkzuuropstapeling.
Osmoregulatie en excretie
Cellen zijn continu onderhevig aan diffusie en osmose (selectieve permeabiliteit vh plasmalemma)
Protozoa in zoetwater ( hypotonisch milieu): cte inwaartse watertransitie. Hebben geen celwand
zoals plantencellen dus opzwellen en ontploffen eencelligen hebben pulserende vacuole die het
osmotisch nr binnengekomen water nr buiten pompt. Dit is een E-vereisend proces.
Meercelligen hebben dit probleem ook: hypertonisch tgover milieu massaal water opnemen
belang vd nieren (functioneel analoog met pulserende vacuole)
Cellen produceren afvalstoffen. Indien toxisch moeten ze snel verwijderd worden. Excretie:
verwijdering van vooral N-houdende afvalstoffen van het metabolisme.
Individuele cellen via diffusie (enkel vr opgeloste stoffen). Bij eencelligen afvalproducten
opgelost in het overtollige water en via pulserende vacuole verwijderd. Bij dieren zorgen de
nieren ervoor dat de concentratie aan afvalstoffen in het bloed niet te hoog wordt.
Gasuitwisseling
O2 moet aangevoerd worden vr de cellulaire ademhaling. O 2 is 1% oplosbaar in water. Uitwisseling
gebeurt via diffusie, geïnitieerd dr de zuurstofgradiënt. Verwijdering van de geproduceerde CO 2
gebeurt analoog. Zolang CO2, dat met water reageert ter vorming van koolzuur, rechtstreeks met
de buitenwereld kan uitgewisseld worden is er geen probleem. Wanneer CO 2 aan het bloed
afgegeven wordt zijn er maatregelen die verzuring verhinderen (zie later).
Prikkelbaarheid en reactievermogen
Op het celoppervlak zijn receptoren vr bepaalde prikkels die prikkels vertalen in een bepaald
gedrag. Ook bij meercelligen zijn individuele cellen prikkelbaar. Bij meercellige dieren is er
daarbij nog de mogelijkheid tot snelle interactie via het zenuwstelsel.
Groei en reproductie
Metabolisme kan resulteren in groei. Wanneer een cel een bepaald volume overschrijdt gaat ze
delen.
Bij prokaryoten wordt vooraf hun genotofoor verdubbeld, dan volgt uitsplitsing van beide
dochtergenotoforen en de overige celinhoud wordt over beide dochtercellen verdeeld. De
genetische informatie wordt onveranderlijk doorgegeven. Bacteriële vermenigvuldiging = binaire
splijting.
Bij eukaryoten is het doorgeven van genetische informatie ingewikkelder omdat hun genoom
uitgesplitst is over chromosomen en ze een functioneel gecompartimentaliseerde celinhoud
hebben. Bij eukaryoten is er een specifiek proces: kerndeling (karyokinese) en celdeling
(cytokinese)
Celcyclus
G1 G0
G1 controlepunt
Mitose
S
G2 controlepunt
G2
Tussen twee mitosen:
INTERFASE: tijdens deze fase
neemt de cel effectief toe in
volume.
Duur vd mitose is vrij constant (bij zoogdieren ± 1u)
G1: de pas gevormde dochtercellen gaan een aantal essentiële organellen bijmaken
de cel gaat haar normale functie uitoefenen (transcriptie, translatie)
duur is zeer variabel en bepalend vr duur vd cyclus
G0: gedifferentieerde cellen die geen deling meer ondergaan blijven in deze fase
G1 controlepunt: bepaalt of cel klaar is vr S fase
S: semi-conservatieve replicatie (DNA verdubbeling)
G2 controlepunt: bevestiging of DNA verdubbeld is
G2: voorbereiding op mitose
chromatine gecondenseerd tot chromosomen
centrosoom wordt verdubbeld
Mitose
Profase:
Verdere condensatie van chromatinedraden tot chromosomen
Elk chromosoom ontdubbeld in twee dochterchromatiden samengehouden dr het centromeer
Centrosomen migreren nr beide polen en vormen de spoelfiguur
Einde als kernmembraan uiteen valt en microtubuli vd spoelfig nr de chromosomen groeien
Microtubuli die nr de chromosomen toe groeien: kinetochoormicrotubuli, nr de pool:
poolmicrotubuli
Metafase:
Begint wanneer de chromosomen zich in het evenaarsvlak bevinden
Dochterchromatiden zijn nr tegengestelde pool gericht
Dit is het ideale beeld om het aantal chromosomen te tellen
Anafase:
Centromeren worden proteolytisch verbroken
Kinetochoormicrotubuli verkorten en poolmicrotubuli worden langer
Dochterchromatiden gaan uit elkaar
Twee identieke sets van chromosomen worden gescheiden = segregatie
Telofase:
Dochterchromatiden hebben pool bereikt
Reassemblage vd kernmembraan
Chromosomen despiraliseren tot chromatine
Cytokinese treedt op: bij diercellen wordt via werking vh cytoskelet de celmembraan van
binnenuit tss beide kernen getrokken
Voor 1-celligen: mitose = aseksuele voortplanting
Genetische informatie wordt onveranderlijk doorgegeven, geen genetische variabiliteit = KLOON
minder overlevingskansen
Pantoffeldiertje: conjugatie
2 pantoffeldiertjes tg elkaar aan
hebben micro en macronucleus
macro: vegetatieve kern
micro: generatieve kern (komt tss bij conjugatie)
ondergaat meiotische delingen (reductiedelingen)
kernen worden uitgewisseld en versmelten opnieuw
= bevruchting (weer oorspronkelijke hoeveelheid
genetische informatie
genetische informatie is uitgewisseld
individuen weer uit elkaar met elk andere
genetische informatie
Plasmodium: mogelijkheid tot vorming van haploïde gameten en sexuele reproductie