Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Voorwoord
Dit is het overzicht van de studiestof voor De ontwikkeling van blok 1.1.1. Het betreft
hier een overzicht van de verplichte literatuur.
Dit overzicht is geschreven naar eigen inzicht van de auteur. Bij het maken van deze
overzichten wordt geprobeerd de kwaliteit zo veel mogelijk te waarborgen.
SlimStuderen.nl kan echter geen verantwoordelijkheid aanvaarden voor het gebruik
ervan. Dit overzicht dient als aanvulling/hulpmiddel en niet ter vervanging van de
verplichte leerstof.
Uiteraard is nadruk verboden. Als je wilt dat wij in staat blijven de verslagen
aan jullie aan te bieden, geef dit verslag dan niet aan derden. Laat hen zelf een
exemplaar aanschaffen!
SlimMededelingen:
Kwaliteit
Om de kwaliteit van de verslagen op een hoog niveau te houden, zijn wij onder
andere afhankelijk van jullie feedback. Heb je opmerkingen, tips of
verbeterpunten? Mail ze dan naar [email protected], dan kunnen we met deze
feedback onze verslagen verder verbeteren.
Auteurs
SlimStuderen is altijd op zoek naar auteurs! Stuur je motivatie en cv naar
[email protected] als je interesse hebt!
Bestellen
Omdat er nog vaak vragen zijn over de levertijd en bestelprocedure, zullen we
deze hier uiteenzetten:
Bestel je middels een éénmalige machtiging, en doe je dit voor 15:00
uur, dan worden de verslagen dezelfde dag verzonden. In principe ontvang
je de verslagen dan de volgende dag, afhankelijk van de bezorging via
PostNL. Het bedrag wordt achteraf éénmalig van je rekening afgeschreven.
Bestel je middels een overschrijving, dan worden de verslagen eenmaal
per week verzonden. Je dient dus zelf het geld over te maken op ons IBAN
Succes met
de tentamens!
(NL81
ABNA 0590 3250 00). Wanneer je gebruik maakt van deze
methode, heb je de verslagen binnen 6 tot 8 werkdagen in huis nadat
het geld op onze rekening staat.
Social media
Bezoek ook eens Facebook.com/SlimStuderen om op de hoogte te blijven van het
verschijnen van nieuwe verslagen en/of supplementen!
Succes met de tentamens!
SlimStuderen.nl
Facebook.com/SlimStuderen
1
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Inhoudsopgave
VOORWOORD .......................................................................................... 1
INHOUDSOPGAVE .................................................................................... 2
A. CELBIOLOGIE .................................................................................... 3
B. GENETICA .......................................................................................... 5
C.
ANATOMIE ....................................................................................... 17
D. HISTOLOGIE .................................................................................... 22
E.
FYSIOLOGIE .................................................................................... 27
F.
PATHOLOGIE ................................................................................... 31
G. RECHT & ETHIEK.............................................................................. 34
Facebook.com/SlimStuderen
2
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
A. CELBIOLOGIE
Alle organismen zijn opgebouwd uit cellen. Dit zijn kleine, door membraan ingesloten,
eenheden gevuld met een geconcentreerde waterige oplossing van chemicaliën en deze
hebben het vermogen om zichzelf te kopiëren door te groeien en zich vervolgens in
tweeën te delen. Cellen kunnen onderling communiceren en zijn de fundamentele
eenheden van het leven.
Eenheid en diversiteit van cellen
Cellen verschillen onderling erg in hun grootte, vorm en chemische eisen. Deze variaties
zorgen ervoor dat de verschillende cellen verschillende functies kunnen uitoefenen.
Sommige cellen zijn gespecialiseerde fabrieken voor de productie van bepaalde stoffen
als hormonen, zetmeel, vet, latex of pigmenten. Anderen zijn motoren, zoals spiercellen
die brandstof verbranden om mechanische arbeid te doen. Enkele van deze cellen zijn zo
gespecialiseerd, dat ze andere cellen nodig hebben om te overleven.
Hoewel verschillende soorten cellen erg verschillen, hebben ze dezelfde basischemie. Ze
bestaan uit dezelfde moleculen, welke aan dezelfde chemische reacties deelnemen.
Verder dragen alle cellen genetische informatie, in de vorm van genen, in DNAmoleculen. Deze informatie is in dezelfde chemische code geschreven, bestaande uit
dezelfde bouwsteentjes en repliceren op dezelfde manier.
Het is geen toeval dat alle cellen zo op elkaar lijken. Alle hedendaagse cellen zijn
ontstaan uit één oercel. Er zit toch verschil tussen de hedendaagse cellen door mutaties.
Deze veranderen het DNA. Mutaties kunnen positief zijn, waardoor ze beter zullen
overleven en reproduceren dan de voorlopercellen, ze kunnen negatief zijn, waardoor ze
minder goed zullen overleven en reproduceren dan de voorlopercellen, of het maakt geen
verschil. Bij seksuele voortplanting ontstaat het nageslacht niet door een deling van een
cel. Het nageslacht ontstaat uit twee cellen, een van elke ouder, waarbij één nieuwe cel
ontstaat. Mutaties en seksuele voortplanting leiden beide tot genetische variatie en
natuurlijke selectie en zijn de basis van evolutie.
Het genoom van een cel is de gehele reeks van nucleotiden in het DNA van een
organisme. Deze geeft de cel instructies over hoe hij zich moet gedragen. Uit een
bevruchte eicel ontstaan vele verschillende soorten cellen zoals spier- en zenuwcellen. Ze
zien er anders uit, maar bevatten toch hetzelfde DNA. Verschillende cellen laten
verschillende genen tot expressie komen, afhankelijk van hun interne toestand en
omgeving.
De prokaryotische en de eukaryotische cel
Organismen die cellen een nucleus/celkern bevatten worden eukaryoten genoemd en
organismen die cellen zonder nucleus bevatten worden prokaryoten genoemd. Onder de
prokaryoten vallen de bacteriën en de archaea (eencellige micro-organismen). De cellen
van de mens worden tot de eukaryotsiche cellen gerekend.
Organellen in de eukaryotische cel
Het belangrijkste organel van de eukaryotische cel is de nucleus. Hierin ligt alle
informatie van de cel opgeslagen. De nucleus bevat DNA-moleculen en is ingesloten door
twee concentrische membranen, de nuclear envelope. Wanneer de cel zich voorbereid
op het delen, wordt het DNA in de kern zichtbaar als individuele chromosomen.
De energievoorziening van een cel wordt geregeld door de mitochondria. Deze
organellen genereren bruikbare energie. Dit doen ze door voedselmoleculen te oxideren
waarbij ATP ontstaat, de algemene chemische brandstof voor de meeste activiteiten voor
de cel. Mitochondria verbruiken zuurstof en geven koolstofdioxide af tijdens deze
activiteit. Dit wordt daarom de cellulaire ademhaling(cellular respiration) genoemd.
Facebook.com/SlimStuderen
3
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Mitochondria bevatten twee aparte membranen, waarvan de binnenste gekronkeld loopt.
Verder bevatten ze hun eigen DNA en reproduceren door zich in tweeën te delen. Om
deze redenen, en omdat mitochondria erg op bacteriën lijken, wordt er gedacht dat
mitochondria afkomstig zijn van bacteriën die ooit een symbiotische relatie zijn
aangegaan met een eukaryotische cel.
Het endoplasmatisch reticulum (ER) is een onregelmatig doolhof van onderling
verbonden ruimtes, ingesloten door een membraan. Hier worden de meeste
celmembraanonderdelen en exportproducten van de cel gemaakt.
Het Golgi-systeem bestaat uit stapels van afgeplatte membraan omsloten schijven.
Deze bewerkt en verpakt ER-moleculen die naar een andere cel of naar een ander
organel moeten worden getransporteerd.
Lysosomen zijn kleine, onregelmatige gevormde organellen. Er vindt intracellulaire
vertering plaats: ze geven voedingsstoffen uit opgenomen voedseldeeltjes vrij en breken
niet-gewenste moleculen af. Deze niet-gewenste moleculen worden gerecycled of
uitgescheiden.
Peroxisomen zijn kleine door membraan ingesloten blaasjes. Ze zorgen voor een veilige
omgeving voor diverse reacties. Waterstofperoxide wordt hierbij gebruikt om toxische
moleculen te inactiveren.
Tussen het ER, het Golgi-systeem en lysosomen vindt voortdurend transport plaats. Dit
transport verloopt via transportblaasjes. Dit gebeurt door middel van endocytose en
exocytose. Exocytose en endocytose worden ook gebruikt om materiaal de cel in of uit
te transporteren.
Het cytoplasma en cytoskelet van de cel
Het cytoplasma is de vloeistof in een cel. Het is een geconcentreerde oplossing van vele
grote en kleine moleculen waar vele chemische reacties plaatsvinden die fundamenteel
zijn voor het bestaan van die cel. Hier wordt de eerste stap gemaakt in het afbreken van
voedselmoleculen en worden de meeste eiwitten gemaakt. Deze productie van eiwitten
wordt gedaan door ribosomen.
Het cytoplasma is wel structureel. Dit komt door het cytoskelet. Dit is een systeem van
eiwitdraden die vaak geankerd zitten aan het plasmamembraan en de nucleus. Er zijn
drie soorten eiwitdraden:
Actinedraden: dit zijn de dunste draden en komen veel in spiercellen voor. Het is
het centrale deel in het mechanisme verantwoordelijk voor de spiercontractie;
Microtubuli: dit zijn de dikste draden en lijken op holle buizen. Ze zijn nauw
betrokken bij de celdeling;
Intermediaire draden: Deze verstevigen de cel.
Aan deze drie draden zitten vaak nog andere eiwitten bevestigd. Samen vormen ze een
systeem van balken, touwen en motoren die de cel zijn mechanische kracht en vorm
geeft en zorgt voor de bewegingen van de cel.
De binnenkant van een cel is constant in beweging. Motoreiwitten (motor proteins)
gebruiken de energie van ATP om organellen en eiwitten door het cytoplasma te dragen.
Facebook.com/SlimStuderen
4
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
B. GENETICA
De bouw van genen
Het genoom beschrijft de combinatie van alle erfelijke factoren. Onder het genoom
wordt één complete set van chromosomen verstaan. De menselijke cellen, somatische
cellen (somatic cells) zijn onder te verdelen in diploïde en haploïde cellen. Haploïde cellen
bevatten 1 set chromosomen, dit zijn 23 chromosomen. Hieronder vallen de gameten, de
geslachtscellen. Diploïde cellen bevatten 2 sets van chromosomen. Deze cellen hebben
23 paar chromosomen, oftewel 46 chromosomen. Ze bevatten dus twee genomen, twee
kopieën van ieder chromosoom. In deze cellen één genoom afkomstig van de vader en
één genoom afkomstig van de moeder.
De chromosomen bestaan uit opgerold DNA. In niet-delende cellen is dit DNA niet
zichtbaar en wordt het chromatine genoemd. Vlak voordat een cel gaat delen,
condenseert het DNA en worden het chromosomen genoemd. Deze chromosomen zijn nu
ook onder de microscoop zichtbaar.
Elk DNA-molecuul is polymeerketen dat wordt opgebouwd uit vier nucleotiden.
Verschillende volgordes van nucleotiden staan voor bepaalde informatie. Deze informatie
kan via transcriptie worden vertaald naar RNA. Deze informatie kan via translatie
worden omgezet naar eiwitten. Deze eiwitten bestaan uit aminozuren. Alle organismen
maken gebruik van dezelfde 20 aminozuren.
DNA
Het DNA-molecuul bestaat uit een pentose suiker, een deoxyribose, een fosfaatgroep en
1 van de vier typen stikstofbasen(nitrogenous bases): cytosine, thymine, adenine en
guanine. In de genetica worden ze aangeduid met hun eerste letter: C, T, A en G.
Cytosine en thymine zijn enkele koolstof-stikstof ringen: de pyrimidines. Adenine en
guanine zijn dubbele koolstof-stikstof ringen: de purines. Een nucleotide is opgebouwd
uit een deoxyribose, een fosfaatgroep en een stikstofbase. De volgorde van de
stikstofbases bepaalt voor welk eiwit een stukje DNA codeert.
DNA vormt een dubbele helix, een gedraaide ladder. De twee zijkanten van de ladder
bestaan uit suiker en de fosfaatgroepen. Tussen de zijkanten lopen treden. Elke tree
bestaat uit twee stikstofbasen, waarvan beide vastzitten aan een andere zijkant. Door
waterstofbruggen worden de twee stikstofbasen bij elkaar gehouden. Cytosine kan 3
waterstofbruggen vormen met guanine. Thymine kan met adenine 2 waterstofbruggen
vormen.
De ruimtelijke bouw van een DNA-molecuul is geen lange rechte lijn. DNA vormt
nucleosomen (nucleosomes) doordat het DNA om histoneiwitkernen (histone protein
cores) wordt gewonden. Deze nucleosomen vormen vervolgens een spiraalvormige
solenoïde. Deze solenoïden zijn opgerold tot het uiteindelijke chromosoom.
Er zijn verschillende soorten DNA. De eerste is het single-copy DNA. Deze reeks DNA
wordt maar één of soms een paar keer gezien in het genoom. Verder is er nog repetitief
DNA. Dit zijn reeksen die erg vaak worden herhaald in het DNA. Dit DNA is onder te
verdelen in twee soorten: het verspreide repetitieve DNA (dispersed repetitive DNA)
en het satelliet DNA (Satellite DNA). Satelliet DNA herhaalt en verzamelt zich in
bepaalde chromosoomgebieden, waarna het eind van het ene stuk, het volgende,
identieke stuk, direct begint. Verspreid repetitief DNA herhaalt zich verspreid in het
genoom.
De celcyclus
Tijdens de ontwikkeling van een individu vindt er continue celdeling plaats. Om deze
celdeling te kunnen laten plaatsvinden doorlopen de cellen continue een bepaalde cyclus
om zich vervolgens te kunnen delen: de celcyclus.
Facebook.com/SlimStuderen
5
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Een cel bevindt zich het grootste gedeelte van zijn leven in de interfase. De interfase is
de periode waarin de cel wordt voorbereid voor de daadwerkelijke celdeling: de mitose.
De interfase bestaat uit drie fases:
G1-fase: de cel groeit, er vindt synthese van RNA en eiwitten plaats;
S-fase: DNA-replicatie;
G2-fase: de cel groeit, reparatie van eventueel beschadigd DNA, de voorbereiding
op de uiteindelijke celdeling.
De duur van de celcyclus varieert per celtype. De M-fase is relatief kort, ongeveer een
uur. De S-fase duurt veel langer. De groeifases, de G1-fase en de G2-fase, kunnen korter
of langer duren, afhankelijk van de vereiste groei die nodig is voor de celdeling. Wanneer
de omstandigheden voor een celdeling erg ongunstig zijn kan de cel in de G0-fase
worden gezet. De cel bevindt zich dan voor een langere periode in een rusttoestand.
Belangrijk bij de celcyclus zijn de CDKSs, de cyclin-dependent kinases. Zij fosforyleren
andere eiwitten waardoor deze geactiveerd of juist geïnactiveerd worden. Ze spelen dus
een belangrijke rol bij de regulatie van de celcyclus. De celcyclus heeft ook een aantal
checkpoints, deze checks worden uitgevoerd door inhibitoren (cyclin dependent kinase
inhibitors). Deze inhibitoren houden fout DNA tegen, dit voorkomt de vorming van
gemuteerde dochtercellen die zich ontwikkelen tot kwaadaardige tumoren.
Vorming spoelfiguur
Microtubuli hechten aan elkaar en via een eiwitstructuur aan de centromeren van de
chromosomen: de kinetochoren. De microtubuli zijn verantwoordelijk voor het uit elkaar
trekken van de gerepliceerde chromosomen. Samen met actine filamenten vormen de
microtubuli een contractiele ring: het mitotische spoelfiguur. De actine filamenten
zitten vast aan de cytoplasmatische kant van de plasmamembraan. Doordat de actine
filamenten en de microtubuli over elkaar glijden, kan het mitotische spoelfiguur
samentrekken. Dit werkt via hetzelfde principe als het samentrekken van spieren. Met
behulp van het mitotische spoelfiguur kan er een plasmadeling optreden: cytokinese.
De M-fase
De M-fase kan worden opgedeeld in de profase, de metafase, de anafase en de
telofase.
Profase condensatie van de gerepliceerde chromosomen. Ze worden nu zichtbaar
onder de microscoop. Buiten de kern wordt een mitotisch spoelfiguur gevormd
vanuit de centrosomen, de polen aan weerszijden van de cel, die uit elkaar gaan.
Buiten de kern vormt het mitotische spoelfiguur. Aan het einde van de profase
wordt de kernenvelop afgebroken. De microtubuli kunnen zo de kinetochoren
bereiken.
Metafase: de chromosomen komen precies tussen de twee polen te liggen.
Anafase: scheiding van de gerepliceerde chromosomen. Elke chromosoom wordt
door het spoelfiguur naar een van de polen getrokken.
Telofase: de chromosomen bereiken de polen van het spoelfiguur. Om de twee
chromosomensets worden nieuwe kernenveloppen gevormd, waardoor twee
nieuwe kern met elk hun eigen materiaal ontstaan. De mitose wordt beëindigd.
Meiose
Wanneer een eicel en een zaadcel bij elkaar komen om een zygote te vormen, worden
hun chromosomen gecombineerd in een enkele cel. Het mechanisme waarbij haploïde
gameten worden gevormd uit een diploïde voorloper heet de meiose. Tijdens de meiose
vinden er twee celdelingen plaats. Elke deling is verdeeld in fases met dezelfde namen
als die van de mitose, maar het proces verschilt soms behoorlijk.
Meiose I
Tijdens de meiose I, de reductiedeling fase (reduction division stage), worden twee
haploïde cellen gevormd uit een diploïde cel. Deze diploïde cellen zijn de oogonia bij
Facebook.com/SlimStuderen
6
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
vrouwen en de spermatogonia bij mannen. Bij meiose II, de equationele deling
(equational division), wordt de haploiïde cel gerepliceerd.
De eerste fase van meiose I is de interfase I, waarbij belangrijke processen zoals de
replicatie van chromosomaal DNA plaatsvindt. De hierop volgende profase I is erg
complex. Het begint met de condensatie van het DNA tot chromosomen. Tijdens de
synapsis vormen de homologe chromosomen paren. Dit gebeurt niet in de mitose.
Wanneer profase I begint, verstrengelen de chromatiden van de twee chromosomen. Elk
paar van verstrengelde homologe chromosomen is bivalent of tetrad. Verder worden
tijdens de profase I chiasmata gevormd, een overkruising van twee chromatiden van
homologe chromosomen. Elk chiasma geeft een punt aan waar de homologe
chromosomen genetisch materiaal uitwisselen. Dit heet crossing over. Er worden
chromosomen gevormd die bestaan uit delen van de originele chromosomen.
De volgende fase is de metafase I. Deze fase is net als bij de mitose gekarakteriseerd
door het afronden van de spoeldraadaanmaak en de opstelling van de bivalenten, die nog
steeds aan de chiasmata vastzitten. De twee centromeren van elk bivalent liggen aan
tegengestelde kanten van het equatoriale vlak.
Tijdens de anafase I verdwijnen de chiasmata en de homologe chromosomen worden
door de spoeldraden naar tegenovergestelde kanten van de cel getrokken. De
centromeren verdubbelen en delen niet, dus er is nog maar de helft van het originele
nummer van chromosomen over.
De volgende fase is de telofase I. Deze begint wanneer de chromosomen de aparte
kanten van de cel hebben bereikt. De chromosomen ontkronkelen licht en een nieuw
kernmembraan begint zich te vormen. De twee dochtercellen bevatten elk een haploïd
aantal chromosomen en elk chromosoom heeft twee zuster chromatiden. Bij mensen
vindt ook de cytokinesis plaats tijdens deze fase. Het cytoplasma is gelijkmatig verdeeld
over de twee dochtercellen in de gameten gevormd bij mannen. Bij vrouwen gaat bijna al
het cytoplasma naar één dochtercel. Deze zal later de eicel worden. De andere dochtercel
zal een poollichaampje worden.
Meiose II
Meiose II begint met de interfase II. Dit is in vergelijking met de Interfase I een korte
fase, er is geen sprake van DNA-replicatie. Profase II lijkt erg op de profase van de
mitose, alleen bevat de nucleus een haploïd aantal chromosomen. Tijdens profase II
worden de chromosomen dichter terwijl ze zich oprollen. Het kernmembraan verdwijnt en
er worden nieuwe spoeldraden gevormd. Dit gaat door in de metafase II. Hier trekken de
spoeldraden de chromosomen in opstelling op het equatoriale vlak. Anafase II volgt. De
centromeren splitsen en elk draagt een enkele chromatide naar een van de kanten van
de cel. De chromatiden zijn gescheiden, maar door de chiasma formatie en de crossing
over, kan het zo zijn dat de nieuwe zusterchromatiden niet identiek zijn.
Telofase II begint wanneer de chromosomen de uiteinden van de cel bereiken. Dan
ontkronkelen ze zich. De nieuwe kernmembranen worden gevormd om elke groep
chromosomen en de cytokinesis verschijnt.
Bij mannen worden alle vier de cellen een spermacel. Bij de vrouw wordt maar 1 van de
vier cellen daadwerkelijk een eicel. De andere drie cellen worden poollichaampjes. De
eerste fase van de oogenesis is al afgerond voordat de vrouw is geboren. De tweede fase
vindt pas plaats na de ovulatie.
DNA-replicatie
Wanneer cellen delen, moeten de dochtercellen hetzelfde DNA bevatten. Hiervoor moet
het DNA eerst verdubbeld worden: DNA-replicatie. De replicatie begint bij een bepaalde
nucleotidevolgorde in het DNA: the origins of replication. De zwakke waterstofbruggen
tussen de stikstofbasen van de DNA-helix worden hier verbroken door het enzym
Facebook.com/SlimStuderen
7
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
helicase, waardoor twee enkele strengen DNA ontstaan. De energie die hiervoor nodig is
wordt gehaald uit de hydrolyse van ATP. Door de opening van het DNA ontstaan er twee
replicatievorken. De 2 vorken bewegen van de origin of replication af in tegengestelde
richting: bidirectioneel. De gevormde vorken zijn asymmetrisch, de ene nieuwe DNAketen wordt gevormd in de 3’-5’ richting, de andere in de 5’-3’ richting.
Het belangrijkste enzym dat betrokken is bij de replicatie van het DNA is DNApolymerase. Het bindt aan de beide replicatievorken en synthetiseert nieuw DNA door
de oude streng als template te gebruiken. DNA-polymerase katalyseert de toevoeging
van een nieuw nucleotide aan het 3’ uiteinde. Dit gebeurt door een fosfodiester binding
tussen het 3’ uiteinde en de 5’-fosfaatgroep te plaatsen.
DNA polymerase kan alleen een nucleotide aan het 3’ uiteinde vastmaken als er al een
DNA-streng is. Het kan niet de vorming van een nieuwe streng starten. Daarvoor zorgt
een RNA-polymerase, welke een RNA-primer vormt van ongeveer 10 nucleotiden,
complementair aan de DNA-streng. Het DNA-polymerase kan vervolgens nucleotiden aan
die RNA-primer toevoegen zodat een complementaire DNA-streng ontstaat.
Het 3’ uiteinde van de beide strengen van het DNA liggen in tegenovergestelde
richtingen. Hierdoor verloopt de replicatie van beide strengen op verschillende manieren.
Bij de leidende streng(leading strand) wordt aan het 3’ uiteinde continu een DNAnucleotide toegevoegd in de richting van de replicatievork. Daar bevindt zich helicase. In
de andere streng is de groeirichting juist van de replicatievork af. Er worden steeds
nieuwe stukjes gesynthetiseerd. Deze nieuwe stukjes worden Okazi-fragmenten
genoemd. Om de replicatie sneller te laten verlopen, worden replicatiesprongen
(replication origins) gemaakt. De replicatie begint dus tegelijk op verschillende punten in
het DNA-molecuul. Deze fragmenten worden Okazaki fragmenten genoemd. De DNAstreng die met behulp van Okazaki fragmenten wordt gerepliceerd heet de lagging
strand.
Om vervolgens een continue DNA-streng te krijgen zijn drie enzymen nodig: nuclease,
repair polymerase en DNA-ligase. Nuclease breekt de RNA-primer af. Repair
polymerase vervangt het RNA door DNA. De okazaki fragmenten worden hierbij als
primer gebruikt. DNA ligase verbindt vervolgens het 5’ einde van het ene fragment met
het 3’ einde van het volgende fragment. Voor deze reactie is de hydrolyse van ATP of
NADH noodzakelijk.
DNA-polymerase kan ook fouten in de DNA-replicatie herkennen en corrigeren. Dit wordt
proofreading genoemd. DNA-polymerase kan dit alleen als het een nieuwe streng in de 5’
naar 3’ richting vormt.
Telomeren zorgen ervoor dat er ook replicatie kan plaatsvinden aan het einde van de
chromosomen. Zonder deze telomeren zouden de DNA-strengen na elke replicatie een
stuk korter worden. Aan het uiteinde van de DNA-streng is namelijk geen plek om een
RNA-primer te plaatsen. Daarom zitten er aan het einde van de DNA-streng speciale
nucleotiden, de telomeren, die het enzym telomerase aantrekken. Dit is een enzym die
kopieën van dezelfde telomeer nucleotiden aan het einde van het chromosoom plakt.
Hierdoor ontstaat een template die het mogelijk maakt de replicatie van de lagging
strand af te maken.
Eiwitsynthese
Na de DNA-replicatie in de nucleus, vindt er eiwitsynthese plaats in het cytoplasma. De
genetische informatie in het DNA wordt door middel van messenger-RNA (mRNA) naar
het cytoplasma getransporteerd. Daar wordt het vervolgens gebruikt om de
samenstellingen van de eiwitten te bepalen. Dit gebeurt met transcriptie en translatie.
Om de transcriptie en translatie uit te kunnen voeren moet het DNA eerst worden
omgezet in RNA. RNA lijkt erg op DNA, maar verschilt op een paar punten. Op de plaats
Facebook.com/SlimStuderen
8
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
van deoxyribose zit ribose en thymine is vervangen door uracil (U). Uracil kan net als
thymine een basenpaar vormen met adenine. Een ander verschil is dat RNA meestal een
enkelstrengs molecuul is in plaats van de dubbele helix bij DNA.
Transcriptie
Bij transcriptie wordt messenger RNA (mRNA) uit DNA gevormd. RNA polymerase II
bindt aan een promoter plaats op het DNA, waarna de RNA polymerase een deel van de
DNA-strengen uit elkaar trekt. Aan een van de DNA-strengen worden mRNA nucleotiden
gebonden. Het mRNA-molecuul kan alleen in de 5’ naar 3’ richting worden gemaakt. Dit
heet stroomafwaarts (downstream). De van 3’ naar 5’ richting wordt stroomopwaarts
(upstream) genoemd. De transcriptie gaat door totdat een basegroep, genaamd de
terminatiesequentie, is bereikt. Dichtbij dit punt worden 100 tot 200 adeninebasen, de
poly-A staart (poly-A tail), toegevoegd aan het 3’ uiteinde van het RNA-molecuul. Dit
zorgt ervoor dat het mRNA-molecuul stabieler is. De mRNA-bases die na de poly-A staart
zijn gekoppeld, raken uiteindelijk verloren. Dit mRNA-molecuul wordt het primaire
transcript genoemd.
Bij sommige genen vindt in alle cellen van het lichaam transcriptie plaats. Dit zijn
zogeheten Housekeeping genen en dienen voor het onderhoud en de stofwisseling van
de cel. De meeste genen dienen echter voor specifieke weefsels op specifieke punten in
de tijd. Deze genen worden alleen in specifieke cellen van de specifieke weefsels
afgelezen.
Veel verschillende eiwitten nemen deel aan de transcriptie. Sommige van deze eiwitten,
de algemene transcriptiefactoren, zijn nodig bij de transcriptie van alle genen. Andere
eiwitten, de specifieke transcriptiefactoren, hebben gespecialiseerde rollen en worden
alleen bij bepaalde genen op bepaalde momenten in de ontwikkeling geactiveerd.
De transcriptie activiteit van specifieke genen kan sterk worden verhoogd door interactie
met sequenties genaamd enhancers. Zij hebben geen direct contact met de genen,
maar specifieke transcriptiefactoren, de activators, kunnen aan deze sequentie binden.
Co-activators kunnen vervolgens binden aan de activators. Aan de co-activators kan
vervolgens de basale transcriptiefactor (general transcription factor) binden. Deze
helpen bij de herkenning en de binding van de co-activators. Waar enhancers helpen met
het verhogen van de transcriptieactiviteit van genen, zorgen silencers voor het
onderdrukken van de transcriptie van genen met behulp van hetzelfde soort interacties.
Specifieke DNA-reeksen worden gelokaliseerd door DNA-bindende motieven (DNAbinding motifs). Dit zijn configuraties in het transcriptiefactoreiwit die het toelaten om
precies en stabiel te passen in een uniek gedeelte van het DNA. Denk hierbij bijvoorbeeld
aan de β-sheet.
De activiteit van de genen kan ook gerelateerd zijn aan een opgerolde vorm of
verstevigde patronen van chromatine. Open chromatine regio’s worden euchromatine
genoemd. Deze zijn gekarakteriseerd door histonacetylering, het bevestigen van
acetylgroepen aan lysineresiduen in de histonen. Acetylering van histonen vermindert
hun binding met DNA. Dit helpt om de chromatine te decondenseren zodat het
toegankelijker is voor de transcriptiefactoren. Euchromatine is dus transcriptioneel actief.
Heterochromatin is daarentegen minder geacetyleerd, gecondenceerder en
transcriptioneel inactief.
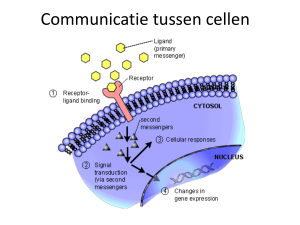
Ook kan de expressie van de genen worden beïnvloed door microRNAs (miRNA). Dit
zijn kleine RNA moleculen, waar geen translatie plaatsvindt. Zij kunnen binden aan
specifieke mRNA reeksen en de expressieniveaus van deze reeksen verminderen.
Transcriptie
Met transcriptie wordt het ombouwen van DNA tot mRNA bedoeld. Een DNA-molecuul
bestaat uit twee strengen, een coderende streng en een matrijs (volgende streng). De
Facebook.com/SlimStuderen
9
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
matrijs wordt gebruikt om het mRNA te maken. Bij deze productie is RNA-polymerase erg
belangrijk.
Als eerste wordt de dubbele DNA-streng geopend. Hierdoor kan RNA-polymerase aan een
binden aan een bepaald punt van de matrijs en voegt daar nucleotiden toe. De plek van
de binding hangt af van de promotor. Bij de promotor zitten transcriptiefactoren, waar
polymerase zich makkelijker kan binden. Hierdoor zal de polymerase zich dus bij de
promotor binden. Dit hele proces gebeurt in de nucleus.
Er is een RNA-streng ontstaan met daarin intronen (introns) en exonen (exons).
Extronen bevatten informatie over de eiwitten die moeten worden gemaakt, het
coderende DNA. De functie van intronen is niet bekend, niet coderend DNA. Deze
intronen moeten uit het RNA worden geknipt. Dit wordt splicing genoemd. De intronen
vormen een soort lusjes, waardoor de exonen met elkaar verbonden zijn. De intronen
worden weggeknipt en er ontstaat een RNA-streng met alleen extronen. Dit overgebleven
stuk mRNA zal naar het cytoplasma migreren. Hiervoor moeten eerst nog een G-kop en
een poly-A-staart worden toegevoegd. Door de G-kop kan het molecuul door het
kernmembraan komen en de poly-A-staart remt de afbraak van het molecuul in het
cytoplasma.
Translatie
Eiwitten worden samengesteld uit een of meer polypeptiden, welke bestaan uit reeksen
aminozuren. In het lichaam komen 20 verschillende aminozuren voor en vier
verschillende RNA-basen. Drie achtereenvolgende RNA-basen staan samen voor één
aminozuur. Een dergelijk triplet heet een codon. Er zijn 64 verschillende codons
mogelijk, maar er bestaan maar 20 aminozuren. Verschillende codons kunnen namelijk
voor hetzelfde aminozuur coderen. Bovendien zijn er drie codons die niet voor een
aminozuur coderen. Dit zijn zogenaamde stopcodons.
Translatie is het proces waarbij het mRNA wordt vertaald naar een polypeptide. De
verschillende codons van het mRNA geven hierbij aan in welke volgorde de aminozuren
moeten worden ingebouwd. Het mRNA kan niet direct binden aan aminozuren, maar
communiceert met moleculen van tRNA (transfer RNA). Elk tRNA-molecuul heeft aan
het 3’ uiteinde een plaats waar een specifiek aminozuur kan hechten. Aan de andere kant
van het molecuul zit een reeks van drie nucleotiden, het anticodon. De drie nucleotiden
van het anticodon binden aan drie nucleotiden van het mRNA. Het bijgevoegde
aminozuur wordt dan overgebracht naar de polypeptideketen en gesynthetiseerd.
De eiwitsynthese vindt plaats in het ribosoom, die bestaat uit enzymatische eiwitten en
ribosomaal RNA (rRNA). Het rRNA helpt het mRNA en het tRNA aan het ribosoom te
binden. Tijdens de translatie bindt het ribosoom eerst aan het startcodon methionine op
het mRNA. Het ribosoom bindt vervolgens het tRNA aan zijn oppervlakte, zodat het paren
van de basen van het tRNA en mRNA kan gebeuren. Het ribosoom beweegt langs de
mRNA-reeks en zodra het ribosoom een stopcodon tegenkomt, stopt de translatie en is
het polypeptide gevormd. Vervolgens wordt het polypeptide vrijgelaten in het
cytoplasma.
Mutaties
Genetische variatie ontstaat onder andere door mutaties. Deze mutaties kunnen in alle
cellen voorkomen. Mutaties in somatische cellen (somatic cells), dit zijn alle cellen
behalve de gameten, kunnen leiden tot kanker. Mutaties in gameetproducerende cellen
(germline cells) leiden tot nieuwe kenmerken in volgende generaties.
Door mutaties kan een verschil tussen individuen ontstaan in een DNA reeks. Deze
variabele reeksen worden allelen genoemd. Allelen zijn bepaalde varianten van een hen.
De locatie van een gen wordt de locus genoemd. Wanneer een persoon twee dezelfde
allelen heeft op beide chromosomen, is de persoon homozygoot. Wanneer er sprake is
Facebook.com/SlimStuderen
10
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
van verschillende allelen, is de persoon heterozygoot. De allelen die aanwezig zijn op een
bepaalde locus is het genotype van het individu. Bij het beschrijven van het genotype
van een individu worden allelen aangegeven met hoofdletters en kleine letters. Een
hoofdletter geeft een dominant allel aan, een kleine letter een recessief allel. Wanneer
het genotype heterozygoot is, zal de eigenschap van het dominante allel domineren over
de eigenschap van het recessieve allel. Wanneer het wel leidt tot een ander aminozuur
zijn twee typen mutaties te onderscheiden: de missense-mutatie en een nonsensemutatie. Bij een missense-mutatie verandert 1 aminozuur. De polypeptide-keten kan
hierdoor anders worden gevouwen, wat kan leiden tot een totaal ander eiwit. Bij een
nonsense-mutatie leidt de mutatie tot een stopcodon. De polypeptide-keten wordt
hierdoor korter.
Ook kunnen er mutaties optreden waarbij 1 of meer baseparen worden verwijderd of
juist worden toegevoegd. Deze mutaties zijn vaak schadelijk en vaak ernstiger wanneer
het aantal baseparen dat verwijderd of toegevoegd is niet deelbaar is door drie. Dit
omdat codons bestaan uit drie baseparen. Wanneer het aantal baseparen dat toegevoegd
of verwijderd is niet deelbaar is door drie, is er sprake van een frameshift mutatie. Bij
dit type mutatie wordt het genetische leesraam verschoven. Het product van het
gemuteerde gen zal geheel veranderd zijn en meestal geen functie meer kunnen
vervullen.
Andere typen mutaties kunnen de regulatie van de transcriptie of translatie veranderen.
Een promotermutatie kan de affiniteit verminderen van de RNA-polymerase voor een
promoterplaats. Dit leidt vaak tot een vermindering van de productie van mRNA en dus
een vermindering van de productie van eiwitten.
Mutaties kunnen ook de splicing van intronen verhinderen wanneer volwassen mRNA is
gevormd uit het primaire mRNA-transcript. Deze zogenoemde Splice-site mutaties
vinden plaats op intron-exongrenzen en veranderen het splicingsignaal dat nodig is om
een intron te verwijderen. Hierdoor wordt het mRNA verkeerd gespliced wat van invloed
is op de uiteindelijke eiwit synthese.
Mutaties kunnen zowel een winst van functie als een verlies van functie van een
eiwitproduct veroorzaken. Winst-van-functiemutaties leiden tot een helemaal nieuw
eiwitproduct,tot een overexpressie of een ongebruikelijke expressie van een bestaand
eiwitproduct. Verlies-van-functiemutaties zijn vaak te zien in recessieve ziektes. Bij zulke
ziektes raakt 50% van het eiwitproduct verloren, maar de andere 50% werkt normaal.
Genetische stoornissen van het menselijke hemoglobine zijn de meest voorkomende
groep van enkel-genziektes. Een voorbeeld hiervan is de sikkelcelziekte. Het wordt
veroorzaakt door een single missense mutatie dat zorgt voor een veranderd aminozuur in
de β-globine keten.
Veel middelen gerelateerd aan de omgeving zoals bijvoorbeeld bestraling kunnen
geïnduceerde mutaties veroorzaken. Deze middelen worden mutagenen (mutagens)
genoemd. Hiertegenover staan spontane mutaties, die ontstaan tijdens de DNAreplicatie. Vaak worden mutaties weer gerepareerd doordat het enzym nuclease foute
nucleotiden wegknipt en DNA-polymerase vervolgens de juiste weer terugzet.
Polymorfismen
Polymorfismen zijn veranderingen in de nucleotidevolgorde die een dermate grote groep
van de populatie voorkomt, dat niet meer kan worden gesproken over een mutatie. Er
zijn verschillende soorten polymorfismen. VNTRs (variable number of tandem repeats)
zijn polymofismen als gevolg van wisselende aantallen van microsatellietherhalingen in
een bepaald DNA-gebied. STRPs(short tandem repeat polymorphisms) zijn een
vergelijkbaar type polymorfismen als gevolg van wisselende aantallen van kleinere
microsatelliet herhalingen. Het soort polymorfisme dat het meeste voorkomt in het
menselijke genoom, bestaat uit varianten van enkele nucleotideposities op een
Facebook.com/SlimStuderen
11
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
chromosoom. Dit zijn SNPs (single nucleotide polymorphisms). Een ander soort
polymorfisme is de CNV (copy number variants). CNVs bestaan uit verschillen in het
aantal herhaalde DNA-reeksen langer dan 1000 baseparen.
Mitochondria
Mitochondria, welke ATP produceren, bevatten hun eigen dubbelstrengs DNA. Ook fouten
in dit DNA kan tot aandoeningen leiden. Het mitochondriale genoom codeert voor twee
rRNAs, 22 tRNAs en 13 polypeptiden die bij de oxidatieve fosforylering betrokken zijn.
Mitochondriaal DNA wordt maternaal overgeërfd en de kans op mutaties is er groot. Van
een aantal ziektes is bekend dat ze veroorzaakt worden door mutaties in het
mitochondriaal DNA.
Onderzoekstechnieken genvariatie
Er zijn verschillende onderzoekstechnieken om genetische variatie te onderzoeken. Dit
kan bijvoorbeeld met bloedgroepen, waarvan het ABO- en Rh-systeem voorbeelden zijn.
Bloedgroepen zijn echter erg beperkt.
Een andere manier om genetische variatie te onderzoeken is eiwitelektroforese
(protein electrophoresis). Door deze techniek is het aantal detecteerbare polymorfe
systemen erg toegenomen. Deze methode maakt gebruik van het feit dat één
verandering van aminozuur binnen een eiwit een lichte verandering in de elektrische
lading van het eiwit kan veroorzaken. Eiwitten met deze lichte verandering in hun
aminozuurreeks zullen anders migreren door een elektrisch geladen gel. De ene kant van
de gel is positief geladen en de andere kant negatief geladen. Afhankelijk van de lading
van het eiwit zal het dus dichter bij de positief geladen of dichter bij de negatief geladen
kant terechtkomen.
Ook kan variatie op DNA-niveau worden gevonden. Technieken die hiervoor gebruikt
worden zijn: southern blotting en restrictiefragmentanalyse. Hierbij wordt gebruik
gemaakt van bacteriële enzymen. Deze enzymen splijten specifieke reeksen van
menselijk DNA. Deze reeksen worden de restrictieplaatsen(restriction sites) genoemd.
Zo wordt het DNA in fragmenten geknipt. Deze fragmenten worden vervolgens door
middel van elektroforese gesorteerd op hun lengte. Dan worden ze overgedragen naar
een solide membraan (Southern blotting), waar ze worden gevisualiseerd door gelabelde
probes te gebruiken. Met dit proces kunnen verwijderingen of verdubbelingen van DNA
worden gevonden, net zoals polymorfismen in de restrictieplaatsen.
DNA moleculen zijn erg klein, dus moeten er erg veel kopieën worden gemaakt om DNA
variatie zichtbaar te maken. Hiervoor wordt het proces PCR (polymerase chain reaction)
gebruikt. Voor PCR zijn twee primers, DNA-polymerase, veel vrije nucleotiden en
genomisch DNA van een individu nodig. Het DNA wordt eerst sterk verhit tot 95 ⁰C.
Hierdoor raken de strengen uit elkaar. Dit proces wordt denaturatie genoemd.
Vervolgens wordt het afgekoeld tot ongeveer 55 ⁰C. Het DNA wordt nu blootgesteld aan
hoge aantallen enkelstrengse primers. welke hechten aan passende stukken DNA. Het
DNA wordt daarna weer verhit tot 72 ⁰C en in aanwezigheid van de vrije stikstofbasen
wordt een complementaire DNA-streng gevormd door DNA-polymerase.
Het nieuw gevormde dubbelstrengs DNA wordt hierna opnieuw verhit. De twee strengen
denatureren vervolgens weer. Het hele proces kan zo herhaald worden, waarbij het
aantal DNA-strengen exponentieel toeneemt.
Een primair doel van vele genetische studies is om de feitelijke reeks van DNA
basenparen waaruit een gen of een deel van een gen bestaat te bepalen. DNAsequencing is een methode die hiervoor wordt gebruikt. Hierbij wordt dideoxy
methode (dideoxy method) gebruikt. Er wordt gebruikt gemaakt van
kettingbeëindigende dideoxynucleotiden. Deze lijken erg op de gewone
deoxynucleotiden, maar missen een hydroxygroep. Hierdoor kan er na
dideoxynucleotide geen deoxynucleotide meer worden ingebouwd. Op deze manier
Facebook.com/SlimStuderen
12
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
worden kettingen gevormd met verschillende lengtes. Deze DNA-fragmenten worden
vervolgens met behulp van een gel-elektroforese gescheiden op lengte. Zo kunnen stap
voor stap de exacte plaatsen van de nucleotiden bepaald. Alle vier de deoxynucleotiden
kunnen worden ingebouwd, die corresponderen met de verschillende nucleotiden: A, T, C
en G.
Het enkelstrengs DNA waarvan de sequentie van moet worden bepaald wordt gemengd
met radioactief gelabelde primers, DNA-polymerase, normale nucleotiden en een van de
soorten deoxynucleotiden. Op iedere positie kan een normale nucleotide worden
ingebouwd, als ook een deoxynucleotide. Dit gebeurt puur random. Afhankelijk van het
inbouwen van een deoxynucleotide of een normaal nucleotidem, wordt de keten wel of
niet beëindigd. Deze methode is erg langzaam, maar door gebruik te maken van
automatische DNA-sequentie, waarbij fluorescerende etiketten worden gebruikt, wordt
het proces versneld.
Mutaties of polymorfismen detecteren in DNA-reeksen is vaak een kritieke stap in het
begrijpen van hoe bepaalde genen specifieke ziektes veroorzaken. Bij het opsporen van
mutaties en bij vele andere toepassingen wordt ook vaak gebruik gemaakt van
microarrays, oftewel DNA chips. Deze DNA-chip heeft honderden spots, waar op de
bodem een bepaald gen is vastgehecht. Door zowel een controlemonster als een
proefmonster aan deze spots toe te voegen, kan gekeken worden of bepaalde genen
aanwezig zijn op het DNA. Het DNA zal dan namelijk binden aan de genen in de spots.
Dit kan zichtbaar worden gemaakt met behulp van fluorescentie.
Populatievariatie
Het voorkomen van veel genetische ziektes verschilt aannemelijk tussen verschillende
populaties. Deze populatievariatie is te begrijpen door de concepten genotypefrequentie
en genenfrequentie. Onder simpele omstandigheden kunnen deze frequenties geschat
worden door individuen te tellen, maar dit kan niet altijd. Hierbij helpt het HardyWeinberg principe. Deze specificeert de relatie tussen genfrequenties en genotypefrequenties en geldt alleen onder panmixia, willekeurige paring. Er geldt: p + q = 1,
waarbij p en q staan voor bepaalde allelen.
Genetische variatie wordt veroorzaakt door natuurlijke selectie (natural selection). Het
verhoogt de populatiefrequentie van gunstige mutaties en verlaagt de populatiefrequentie van ongunstige mutaties in een bepaalde omgeving. Een andere kracht die
ziektegenen laat variëren in frequentie in populaties is de genetische drift (genetic
drift). Genetische drift is een willekeurig evolutionair proces dat grotere veranderingen in
genfrequenties produceert in kleinere populaties. Genen worden bij toeval verspreid,
waardoor de frequentie van een allel binnen een populatie kan toenemen of afnemen.
Het stichtereffect (founder effect) is een gespecialiseerde versie van de genetische
drift. Hierin kunnen kleine stichterpopulaties door hun kleine omvang grote
veranderingen in genfrequentie creëren, waarna ook relatief grote populaties enkele
drifteffecten ervaren. Gene flow vindt plaats wanneer populaties migranten uitwisselen
die met elkaar paren. De nakomelingen zullen steeds meer op elkaar gaan lijken.
Genotype, fenotype en overerving
George Mendel heeft twee principes naar voren laten komen in zijn werk. De eerste is het
principe van scheiding(principle of segregation), waarin staat dat seksueel
voortplantende organismes genen bezitten die voorkomen in paren en dat slechts één lid
van dit paar aan het nageslacht wordt doorgegeven. Dit kan worden verklaard door de
haploïde geslachtscellen.
Zijn tweede principe is het principe van onafhankelijke assortiment (principle of
independent assortment). Dit principe stelt dat genen op verschillende loci onafhankelijk
worden overgebracht.
Facebook.com/SlimStuderen
13
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Het genotype is gedefinieerd als een genetische samenstelling van een individu op een
bepaalde locus. Het fenotype (phenotype) is wat er daadwerkelijk lichamelijk of klinisch
wordt waargenomen. Individuen met verschillende genotypen, een homozygoot genotype
en een heterozygoot genotype, kunnen hetzelfde fenotype hebben. Het fenotype wordt
naast het genotype ook bepaald door omgevingsfactoren
Pedigree
De pedigree is een van de meest gebruikte tools in de medische genetica. Het laat de
relaties tussen familieleden zien en laat zien welke familieleden wel of niet aangetast zijn
door een genetische ziekte. Een pijl geeft de proband aan, het eerste lid van de familie
waarbij de diagnose voor een ziekte is gesteld. De proband wordt soms ook het index
geval (index case) of propositus/proposita genoemd.
Autosomaal vs. recessief
Autosomale dominante overerving wordt gekarakteriseerd door verticale transmissie van
het ziektefenotype, vrijwel alle generaties zijn aangedaan en zowel mannen als vrouwen
worden aangedaan. De overerving verloopt immers niet via de geslachtscellen.
De waarschijnlijkheid dat een individuele nakomeling getroffen wordt door een ziekte
wordt het herhalingsrisico (recurrence risk) genoemd. Het herhalingsrisico voor een
autosomaal dominante stoornis is 50%.
Autosomale recessieve overerving wordt gekarakteriseerd door clustering van het
ziektefenotype tussen broers en zussen, maar de ziekte wordt meestal niet gezien bij
ouders of andere voorouders. Meestal worden gelijke aantallen van getroffen mannen en
vrouwen gezien en bloedverwantschap van de ouders kan aanwezig zijn. Het
herhalingsrisico voor autosomale recessieve ziektes is normaal gesproken 25%.
Quasidominante overerving, met een herhalingsrisico van 50%, wordt gezien wanneer
een getroffen homozygoot en een heterozygoot samen voor nakomelingen zorgen.
Hoewel het onderscheid tussen dominante en recessieve aandoeningen niet rigide is, zal
een dominant allel wel een ziekte produceren in een heterozygoot, terwijl een recessief
allel dit niet zal doen. In sommige gevallen kan een ziekte erfelijk zijn in zowel
autosomaal dominante als in autosomaal recessieve wijze, afhankelijk van de aard van
de mutatie die het genproduct wijzigt.
De meeste genetische ziektes variëren in de mate van expressie. In sommige gevallen
heeft een individu wel een ziekte gecorreleerd genotype, maar niet het bijbehorende
fenotype. Ook worden soms bij individuen genetische ziektes gevonden, die tot dan toe
niet in de familie voorkwamen. Er zijn verschillende oorzaken die dit kunnen verklaren.
Zo kan er een nieuwe mutatie optreden, waardoor een genetische ziekte wordt
veroorzaakt. Het herhalingsrisico voor broers en zussen is dan erg klein, maar het
herhalingsrisico voor de nakomelingen van de persoon kunnen substantieel verhoogd
zijn.
Kiembaan mozaïcisme (Germline mosaicism) ontstaat als de gehele of een deel van de
kiembaan van een ouder wordt aangetast door een mutatie, maar de somatische cellen
niet. Het verhoogt het herhalingsrisico voor nakomelingen van de mozaïekouder.
Geslachtsgebonden overerving
X-gebonden recessieve ziektes komen meer bij mannen voor dan bij vrouwen. Dit komt
doordat vrouwen twee kopieën van het X-chromosoom hebben en mannen maar één.
Mannen zijn dus bij één ziekteallel ziek, terwijl vrouwen er twee moeten hebben om ziek
te worden. X-gebonden recessieve overerving is gekarakteriseerd door een absentie van
vader-op-zoon overerving, overgeslagen generaties wanneer genen doorgegeven worden
via vrouwelijke dragers en een overwicht van getroffen mannen. Het herhalingsrisico van
X-gebonden recessieve stoornissen zijn is dan voor autosomale stoornissen. Het risico
hangt af van het genotype van beide ouders en het geslacht van de nakomeling.
Facebook.com/SlimStuderen
14
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
X-inactivatie is een willekeurig proces en daarom ervaren sommige vrouwelijke
heterozygoten inactivatie van de meeste normale X-chromosomen in hun cellen. Deze
manifesterende heterozygoten(manifesting heterozygotes) zijn meestal licht
getroffen.
X-gebonden dominante aandoeningen vertonen karakteristieke patronen van overerving.
Ze komen twee keer zo vaak voor bij vrouwen als bij mannen, overgeslagen generaties
zijn ongewoon en vader-op-zoon overerving bestaat niet.
Het Y-chromosoom bevat enkele tientallen Y-gebonden, of holandrische (holandric),
genen. Deze genen kunnen alleen van vader op zoon worden doorgegeven.
Cytogenetica
Ziektes ontstaan niet alleen door fouten in enkele genen. De studie die zich bezig houdt
met chromosomen en hun abnormaliteiten heet de cytogenetica.
Ons vermogen om chromosomen te bestuderen is sterk verbeterd door de tijd heen.
Vanaf het midden van de 19e eeuw kunnen we chromosomen tijdens de metafase door
middel van een microscoop zien. Aan het begin van de jaren ’50 van de vorige eeuw
kwamen er enkele betere technieken bij. Door het gebruik van een hypotonische
oplossing, zwellen de cellen op, breken de nuclei en raken de individuele chromosomen
beter gescheiden. Ze zijn dus apart te zien. Ook worden kleurende materialen gebruikt,
die verschillend geabsorbeerd worden door verschillende delen van chromosomen en
daarmee karakteristieke lichte en donkere banden creëren. Deze banden helpen om
individuele chromosomen te identificeren.
In een karyotype of karyogram worden chromosomen geordend naar lengte.
Chromosomen kunnen hierbij aan de hand van de positie van het centromeer in drie
groepen worden ingedeeld. Een Acrocentrisch (acrocentric) chromosoom heeft zijn
centromeer bij het uiteinde, waar het telomeer (telomere) zit. Submetacentrische
(submetacentric) chromosomen hebben hun centromeer ergens tussen het uiteinde en
het midden van het chromosoom en metacentrische (metacentric) chromosomen
hebben hun centromeer mooi in het midden van het chromosoom.
Behalve genen, kunnen ook hele of gedeeltelijke chromosomen gemuteerd raken. Er zijn
verschillende technieken waarmee chromosoomfouten kunnen worden opgespoord.
Voorbeelden hiervan zijn FISH en CGH.
Cellen die een veelvoud van 23 chromosomen hebben, worden euploïd genoemd.
Triploïde (69 chromosomen) en tetraploïde (92 chromosomen) cellen zijn gevonden bij
de mens. De meeste polypoïde cellen worden spontaan afgebroken en ze zijn allemaal
niet in staat om op de lange termijn te overleven.
Cellen die missende of toegevoegde individuele chromosomen bevatten worden
aneuploïd genoemd. De belangrijkste oorzaak van aneuploïdie is nondisjunction.
Hierbij falen de chromosomen om uit elkaar te gaan tijdens de meiose. Dit kan gebeuren
bij zowel meiose I als meiose II. Meestal bevat zo’n aneuploïde cel een monosomie of
een trisomie. Autosomale monosomieën zijn bijna nooit levensvatbaar, maar sommige
autosomale trisomieën kunnen wel overleven. Een voorbeeld van een trisomie die kan
overleven is trisomie 21. Individuen met deze trisomie hebben het Downsyndroom.
Andere levensvatbare trisomieën zijn trisomie 13 en trisomie 18.
Onder de levend geboren baby’s, heeft ongeveer 1 op de 400 jongetjes en 1 op de 650
meisjes een bepaalde vorm van een aneuploïde van de geslachtschromosomen. Deze zijn
vooral het gevolg van X-inactivatie. De gevolgen van deze vormen van aneuploïde zijn
minder ernstig dan die van autosomale aneuploïdes, met uitzondering van de afwezigheid
van een X-chromosoom. Alle aneuploïde geslachtschromosomen zijn in ten minste enkele
gevallen met leven verenigbaar.
Facebook.com/SlimStuderen
15
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Enkele andere chromosoomfouten zijn:
Translocatie: met een translocatie (translocation) wordt de uitwisseling van
genetisch materiaal tussen niet-homologe chromosomen bedoeld;
Contiguous gene syndromes: microscopisch waarneembare
chromosoomdeleties, die zowel bij een terminale deletie (terminal deletion),
waarbij een enkele breking tot verlies van het chromosomenuiteinde leidt, als bij
een interstitiële deletie (interstitial deletion), waarbij twee breuken optreden en
het materiaal tussen de breuken verloren gaat, tasten vaak een groot aantal
genen aan en leiden zo tot herkenbare syndromen;
Subtelomere herschikkingen (subtelomeric rearrangements): betrekken
deleties of duplicaties van DNA in de gen-rijke regio’s in de buurt van telomeren;
Chromosoominversies: ouders hebben meestal een normaal fenotype, maar
kunnen nakomelingen met deleties of duplicaties produceren.
Chromosoomafwijkingen resulteren doorgaans in een vertraging in de ontwikkeling,
mentale retardatie, karakteristieke gelaatstrekken en verschillende vormen van
aangeboren afwijkingen. Ondanks enige overlap van fenotypische kenmerken, kunnen
veel chromosoomafwijkingen worden herkend door klinisch onderzoek. Gebalanceerde
translocatie in somatische cellen kan soms tot maligniteiten leiden door het onderbreken
of wijzigen van genen of hun regulerende reeksen. De syndromen van chromosomale
instabiliteit betrekken allemaal verhoogde frequenties van chromomosoombreuken en
een verhoogd risico op maligniteiten. Ze zijn allemaal geassocieerd met defecten in de
DNA-replicatie of het herstel.
Facebook.com/SlimStuderen
16
Op zoek naar een bijbaan?
Combineer studie en werk! SlimStuderen is op
zoek naar getalenteerde auteurs voor de studies
Bedrijfskunde, IB en Economie aan de Rijkuniversiteit Groningen. Als auteur maak je een
samenvatting van literatuur of colleges voor
een vak naar keuze. De enige bijbaan waarbij je
tijdens je werk met je studie bezig bent!
De voordelen op een rijtje:
Ruime vergoeding
Gratis samenvattingen
Informele werksfeer
Studerend geld verdienen
Spreekt jou dit aan? Stuur dan een e-mail naar
[email protected] en wie weet maak jij
straks deel uit van het leukste team van
Groningen!
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
C. ANATOMIE
Algemene concepten
Er worden in de geneeskunde diverse termen gebruikt om posities in het lichaam te
beschrijven.
Term
anterior
caudaal
craniaal
contra-
Nederlandse betekenis
voor
Beneden
Boven
Tegengesteld
Term
interipsilateraal
mediaal
dexter
distaal
dorsaal
Rechts
verder van de romp af *
aan de kant van de rug
medius
posterior
proximaal
endo-
aan de binnenkant
rostraal
epiinferior
sub-
erop/erover
aan de onderkant
Onder
sinister
superior
ventraal
Nederlandse betekenis
tussen
dezelfde
aan de zijkant
aan de kant van het
midden
in het midden
achter
dichter naar de romp
toe *
aan de kant van het
gezicht **
links
aan de bovenkant
aan de kant van de
buik
*Deze term wordt alleen gebruikt in ledematen
**Deze term wordt alleen gebruikt in hoofd/hals
Om aan te geven hoe er tegen een doorsnede van het lichaam wordt aangekeken, wordt
er onderscheid gemaakt in drie anatomische vlakken.
Term
Planum transversale (axiale)
Planum frontale (coronale)
Planum sagittale
Nederlandse betekenis
Een horizontale doorsnede: deelt het
lichaam op in ‘onder’ en ‘boven’.
Een verticale doorsnede: deelt het lichaam
op in ‘voor’ en ‘achter’.
Een verticale doorsnede: deelt het lichaam
op in ‘links’ en ‘rechts’.
Somatisch (somatic): met betrekking op het lichaam. Wanneer klachten van somatische
aard zijn, zijn het dus lichamelijke klachten.
Visceraal (visceral): met betrekking op de ingewanden. Bijvoorbeeld het viscerale
zenuwstelsel geeft signalen door aan de ingewanden.
Tussen de longen en de borstholte is er een afscheiding gevormd door een bursa
(sereuze zak). Het is een membraan dat dubbel geplooid is. Het longvlies, ook wel
pleura visceralis genoemd, is het deel van het dubbelgevouwen vlies dat aan de longen
grenst. Het borstvlies (pleura parietalis) grenst aan de borstholte. De pleurale ruimte,
de ruimte tussen deze twee membranen, is gevuld met een klein beetje vocht. Dit vocht
maakt het mogelijk dat tijdens de ademhaling de membranen langs elkaar heen kunnen
bewegen. Te veel vocht veroorzaakt bijvoorbeeld longontsteking.
Signalen worden doorgegeven door middel van zenuwen. Er wordt onderscheid gemaakt
tussen drie verschillende soorten zenuwcellen met hun eigen functie.
Sensorische zenuwcel (sensory fiber): deze zenuwcel geeft prikkels vanuit het
lichaam door aan het centrale zenuwstelsel. Het gaat hier om exteroceptieve
prikkels (pijn, aanraking en temperatuur) en proprioceptieve prikkels uit spieren,
pezen en gewrichten.
Facebook.com/SlimStuderen
17
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Motorische zenuwcel (somatic motor fiber): geeft signalen vanuit het centrale
zenuwstelsel door aan spieren.
Viscerale motorische zenuwcel (visceral motor fiber): deze zenuwcellen van
het sympathische deel van het autonoom zenuwstelsel nemen signalen over van
alle zenuwen uit de wervelkolom en brengen signalen naar de gladde spieren van
bloedvaten en klieren en naar de arrector pili spieren van de huid. Arrector pili
spieren zijn de haarspiertjes, die ervoor zorgen dat haren overeind gaan staan
wanneer deze zijn samengetrokken.
Schakelcel: liggen in de hersenen en in het ruggenmerg. Ze zorgen voor
connectie met andere schakelcellen binnen het centrale zenuwstelsel en ze maken
contact met motorische en sensorische neuronen. Hierdoor kan informatie van de
zintuigen het goede gebied in de hersenen bereiken en kan een passende
motorische reactie plaatsvinden.
Een typische rugzenuw komt uit het ruggenmerg vanuit zenuwwortels die samenkomen
tot twee zenuwwortels. De voorste wortel (anterior (ventral) root) bestaat uit
motorische vezels, die zenuwcellichamen in de voorste hoorn van het ruggenmerg de
grijze stof passeren, om effectororganen te bereiken.
De achterste wortel (posterior (dorsal) root) bestaat uit sensorische zenuwen die
zenuwimpulsen uit sensorische receptoren in verschillende delen van het lichaam naar
het centraal zenuwstelsel brengen.
De voorste en achterste wortel komen samen in de tussenwervelschijf om daar een
spinale zenuw te vormen, dit scheidt onmiddellijk in twee rami (takken van zenuwen): in
een posterior ramus en een anterior ramus.
Posterior rami: leveren zenuwvezels aan met slijmvlies beklede gewrichten van
de wervelkolom, diepe spieren van de rug en de overliggende huid.
Anterior rami: leveren zenuwvezels aan voorkant en aan de zijkant gelegen
gebieden in de romp en de ledematen die daaruit voortkomen.
Specifieke/regionale anatomie: wervelkolom en ruggenmerg
De wervelkolom is de basis van het bewegingsapparaat. Aan de binnenkant zitten de
zenuwwortels en het ruggenmerg, en aan de buitenkant vormt het de aanhechting van
het bekken en belangrijke spieren van de romp. De wervelkolom bestaat uit vele wervels,
die allemaal een nummer bevatten.
Bovenin zitten zeven cervicale- (of nek) wervels: C1 t/m C7. Daaronder zitten de twaalf
thoracale- (of borst) wervels: Th1 t/m Th12. De onderste vijf wervels betreffen de
lumbale (of lenden-) wervels: L1 t/m L5. Helemaal onderaan zit het sacrum (heiligbeen)
met het staartbeentje.
Vanaf het hoofd naar de borst lopen zeven cervicale wervels. Eigenschappen van een
typische cervicale wervel zijn:
Een relatief groot foramen vertebrale (tussenwervelgat)
Een ingestulpt, ovaal wervellichaam
Een stompe processus spinosus (doornuitsteeksel)
Binnen de dwarsuitsteeksels een rond foramen transversarium, die de belangrijke
bloedvaten naar de hersenen beschermen.
Wervels kunnen makkelijk bewegen ten opzichte van elkaar; dit komt door de elastische
tussenwervelschijven, maar ook door de gewrichten die de onderliggende en
bovenliggende wervels met
elkaar in verbinding brengen.
Bij voor- of achteraanzicht staat de gehele wervelkolom in een rechte lijn. Bij sommige
mensen is er een lichte kromming naar lateraal, maar als het binnen bepaalde grenzen
blijft wordt er niet gesproken over een pathologische kromming.
Facebook.com/SlimStuderen
18
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
De wervelkolom vertoont in het sagittale vlak vier verschillende krommingen. Van
caudaal naar craniaal zijn dat:
De sacrale kromming; die staat vast doordat de sacrale wervels vergroeid zijn met
elkaar; deze kromming is bol naar voren.
De lumbale lordose; bolvormig naar achteren.
De thoracale kyfose; bolvormig naar achteren.
De cervicale lordose; bolvormig naar achteren.
Wanneer er sprake is van een pathologische kromming, zijn er twee verschillende
vormen. Bij een hyperlordose is de vorm van de rug te hol, en bij een hyperkyfose is de
vorm van de rug te bol. Bij een scolliose is er een onnatuurlijke draaiing van de
wervelkolom. Hierbij hebben de wervels een zijdelingse kromming en/of zijn ze verdraaid
in de lengte-as.
De hersenvliezen bieden bescherming aan de twee belangrijkste organen van het
zenuwstelsel; de hersenen en het ruggenmerg. De hersenvliezen zijn omhulsels voor de
vele zenuwen die door de openingen aan de schedelbasis uittreden in de wervelkolom.
Embryologie
De embryogenese begint zodra een öocyt is bevrucht. De embryogenese bestaat uit een
aantal stadia. De zygote doorloopt de morulafase, de blasulafase en de gastrulatie.
Gastrulatie (gastrulation) is het begin van de morfogenese; de kiemlagen ontstaan.
Morfogenese betekent dat de ontwikkeling van het lichaam begint. Dit proces begint in
de derde week van de zwangerschap. Er zijn drie kiembladen die belangrijk zijn voor de
ontwikkeling van specifieke organen en weefsels.
Embryonaal ectoderm (Embryonic ectoderm): dit is het buitenste kiemblad van
een embryo. Doet de epidermis, het centraal en perifere zenuwstelsel, de ogen,
de inwendige oren en bindweefsels in het hoofd ontstaan;
Embryonaal endoderm (Embryonic endoderm): dit is het binnenste kiemblad
van een embryo. Dit is de bron van de epitheliale bekleding van de luchtwegen en
het spijsverteringsstelsel;
Embryonaal mesoderm (Embryonic mesoderm): dit is het middelste kiemblad
van een embryo. Het is het begin van alle skeletspieren, bloedcellen en de
bekleding van bloedvaten, serosale bekleding van lichaamsholten, de kanalen en
organen van het voortplantings- en uitsscheidingssysteem. Het is de bron van alle
bindweefsels.
Na de gastrulatie vindt de organogenese en de neurulatie plaats. Neurulatie
(neurulation) is het proces dat betrokken is bij de vorming van het centraal zenuwstelsel.
De neurulatie vindt plaats in de vierde week van de zwangerschap. Vanuit het ectoderm
wordt eerst de neurale plaat (neural plate) gevormd. Op de neurale plaat ontwikkelt
zich de primitieve streep (primitive streak). Vanuit de primitieve streep wordt de
neurale groeve en vervolgens de neurale buis gevormd; dit wordt veroorzaakt door
plooiing en migratie van epiblastcellen uit het ectoderm. De hersenen, het ruggenmerg
en de perifere zenuwen komen voort uit de neurale buis.
Vanuit het paraxiale mesoderm ontstaan somieten (somites). Aan het eind van de derde
week van de zwangerschap verandert, concentreert, en begint het paraxiale mesoderm
te scheiden in gepaarde kubusvormige lichaampjes; dit zijn de somieten. Deze blokjes
mesoderm liggen aan elke zijde van de ontwikkelende neurale buis. Somieten vormen de
structuren myotoom, dermatoom en sclerotoom. Vanuit deze structuren worden spieren,
huid en de gewrichten gevormd. Vanuit het intermediaire mesoderm ontstaat het
urinewegstelsel.
Facebook.com/SlimStuderen
19
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Het intraembryonaal coeloom (intraembryonic coelom) scheidt het lateraal mesoderm
in twee lagen:
Een somatische of pariëtale laag van lateraal mesoderm dat ononderbroken is met
het extraembryonaal mesoderm dat het vruchtvlies, de amnion, bedekt.
Een viscerale laag van lateraal mesoderm dat ononderbroken is met het
extraembryonale mesoderm dat het navelstrengblaasje bedekt.
Tijdens de tweede maand van de zwangerschap is het intraembryonaal coeloom gesplitst
in drie lichaamsholten: de pleurale holten, de buikholte en een pericardiale holte (holte
die het hart op de juiste plaats houdt).
De organogenese (organogenesis) is de fase waarin de organen in het lichaam
ontstaan, het is de fase na de neurulatie. Dit vindt plaats in de derde tot achtste week
van de zwangerschap. De kiemlagen veranderen door drie processen: vouwen, splitsen
en condenseren. De cellen van elke kiemlaag migreren, vermenigvuldigen zich en
veranderen in verschillende weefsels die de organen vormen.
Week 3: begin van ontwikkeling van het brein, het hart, de bloedcellen, de
bloedsomloop, het ademhalingsstelsel, het ruggenmerg en de spijsvertering;
Week 4: begin van ontwikkeling van de botten, gezichtsstructuren, ledematen
(aanwezigheid van armen en benen), zenuwweefsel en het spijsverteringskanaal.
Het hart ontwikkelt zich verder en begint te kloppen;
Week 5: begin van ontwikkeling van de ogen, neus, nieren, longen. Ontwikkeling
van het hart, brein, zenuwweefsel en spijsverteringskanaal gaat verder;
Week 6: begin van de ontwikkeling van de handen, voeten, vingers en tenen.
Ontwikkeling van het brein, hart en ademhalingsstelsel gaan door;
Week 7: begin van de ontwikkeling van haarzakjes, tepels, oogleden en
geslachtsorganen. In deze week is de eerste productie van urine in de nieren en
de eerste waarneming van hersengolven;
Week 8: de gelaatstrekken zijn meer uitgesproken, de binnenste organen zijn
ontwikkeld, het brein kan signalen naar spieren sturen om te bewegen en de
buitenste geslachtsorganen beginnen te groeien.
Na deze fase zijn buitenste en binnenste structuren van het lichaam gevormd. Vanaf nu
is de embryonale fase afgesloten en wordt het geen embryo meer genoemd, maar een
foetus.
De cephalocaudale ontwikkeling beschrijft de ontwikkeling van de lichaamscontrole
van een kind van het hoofd naar de lagere delen van het lichaam. Een kind zal
bijvoorbeeld het eerst controle hebben over het hoofd, het bovenlichaam en de armen.
Pas later zal de baby controle krijgen over de been en het onderste deel van de romp.
De laterale ontwikkeling beschrijft dat een kind eerst controle krijgt over zijn of haar
grove motoriek. Pas later volgt de controle over de fijne motoriek.
Ontwikkelingsstoornissen
Neurale buisdefecten zijn aangeboren afwijkingen van de neurale buis. Uit de neurale
buis van een ongeboren kind ontstaan het ruggenmerg en de hersenen. De neurale buis
ontwikkelt zich al op de 22e dag na de bevruchting. De neurale buis is eerst nog open,
maar deze moet binnen een paar dagen dichtgroeien. Op het moment dat dit
dichtgroeien niet goed verloopt is er sprake van een neuraal buisdefect. Er zijn
verschillende soorten neurale buisdefecten, omdat de neurale buis op meerdere plekken
niet goed dicht zou kunnen groeien.
Facebook.com/SlimStuderen
20
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Wanneer het proces van sluiten van de neurale buis niet goed verloopt, levert dit
ernstige problemen op. Wanneer de sluitingsstoornis craniaal is, leidt dit tot
anancefalie. Bij anancefalie blijven de grote hersenen achter in ontwikkeling. Een
caudale sluitingsstoornis leidt tot spina bifida aperta (open ruggetje). Bij spina bifida
aperta zit er een open gedeelte op de rug. Er zijn twee verschillende vormen die kunnen
voorkomen: een meningocèle en myelomeningocèle. Bij een meningocèle bevat het
open gedeelte alleen hersenvliezen of liquor; in 20% van de gevallen komt dit voor. Bij
een myelomeningocèle bevat het open gedeelte ook uittredende zenuwwortels en/of
ruggenmerg; dit komt in 80% van de gevallen voor. Bij spina bifida aperta is er vaak
sprake van hydrocefalie (waterhoofd). Het is mogelijk dat er een aanlegstoornis ter
hoogte van het achterhoofdsgat en de achterste schedelgroeve is (Chiarimalformatie) of
een aanlegstoornis aan de grote hersenen.
Patiënten met een laag lumbale myelomeningocèles kunnen later in veel gevallen, soms
met hulpmiddelen, lopen. Patiënten met een thoracale myelomeningocèle zijn zullen
rolstoel gebonden zijn voor de rest van hun leven.
Bij spina bifida occulta is er een sluitingsdefect van de lumbosacrale bogen
(wervelbogen). Doordat de wervelbogen niet goed gesloten zijn, kunnen er ook
structuren in de zwelling voorkomen buiten de wervelkolom. In de zwelling kan ook een
lipoom (vetbultje) worden aangetroffen.
Spina bifida is een aandoening die door verschillende factoren kan worden veroorzaakt.
Naast het feit dat het erfelijk is, kan het ook door exogene factoren komen. Wanneer de
moeder tijdens de zwangerschap anti-epileptica slikt of als de zwangere obesitas heeft, is
er een verhoogd risico op een kind met spina bifida. Tegenwoordig wordt vrouwen met
een kinderwens aanbevolen om foliumzuur te slikken voor en een deel tijdens hun
zwangerschap; dit verlaagt het risico op een kind met spina bifida. Het aantal kinderen
dat geboren wordt met defecten aan de neurale buis is afgenomen door echo-onderzoek
bij de 20e week in de zwangerschap en het toedienen van foliumzuur.
Wanneer er geen complicaties optreden bij de ontwikkeling van het centrale
zenuwstelsel, verloopt dit proces in de volgende stadia:
In de derde week van de zwangerschap begint de ontwikkeling, waarbij als eerste
de neurale plaat ontstaat. In de plaat ontstaat vervolgens een groeve (de neurale
buis). Het craniale deel van de neurale buis zal zich verder ontwikkelen tot de
hersenen. Uit het caudale deel zal het ruggenmerg ontstaan.
Er vormen primaire hersenblaasjes in het craniale deel van de neurale buis. Dit
worden de voorhersenen, middenhersenen en achterhersenen.
De hersenblaasjes ontwikkelen zich tot vijf secundaire hersenblaasjes. In elk
hersenblaasje zit een neuraal kanaal dat uitgroeit tot een ventrikel. Dit worden
uiteindelijk de hersenkamers/de ventrikels.
De twee hersenhelften worden gevormd, de tussenhersenen splitsen zich en
verschillende onderdelen ontstaan (bijvoorbeeld de hersenzenuwen).
Facebook.com/SlimStuderen
21
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
D. HISTOLOGIE
Weefseltypen
Weefsels zijn groepen cellen die allemaal dezelfde vorm en functie hebben, het zijn
gelijksoortig gedifferentieerde cellen. Er zijn vier verschillende soorten weefsels te
onderscheiden die allemaal hun eigen kenmerken hebben: epitheelweefsel, bindweefsel,
spierweefsel en zenuwweefsel. Tussen de cellen van de verschillende typen weefsels
bevindt zich extracellulair materiaal. Het extracellulaire materiaal zorgt onder andere
voor ondersteuning van de cellen.
Alle weefseltypen komen uiteindelijk voort uit de zygoot, een vaste bal met 2-8 cellen.
De zygoot ontwikkelt zich tot een blastocyste met 50-60 cellen, hierbij ontstaat een
holte met stamcellen. De buitenste laag van deze holte wordt de trofoblast genoemd. Na
8-10 dagen heeft de blastocyste 100 cellen, deze bestaan uit stamcellen, epiblastcellen
en hypoblastcellen. De blastocyste komt na 12-14 dagen in de uterus, waar er een
tweebladige kiemschijf (bilaminar embryonic disc) ontstaat met epiblast- en
hypoblastcellen. De epiblastcellen ontwikkelen zich tot de amnionholte (amniotic cavity),
en de hypoblastcellen ontwikkelen zich tot de dooierzak (yolk sac).De dooierzak is de
voorloper van de placenta.
In de derde week ontstaat er een driebladige kiemschijf (trilaminar embryonic disc). Dit
gebeurt doordat epiblastcellen via de primitieve streep naar het gebied tussen de epiblast
en de hypoblast migreren. Deze cellen ontwikkelen zich tot het mesoderme
kiemweefsel. De overige epiblastcellen ontwikkelen zich tot het ectoderme
kiemweefsel. De hypoblastcellen vormen uiteindelijk het endoderme kiemweefsel.
De vorming van deze drie kiemweefsels wordt de gastrulatie genoemd. Uit deze
kiemweefsels worden uiteindelijk alle weefsels van de embryo gevormd:
Ectoderm: de epidermis, het tandglazuur, de hersenen, het netvlies, de klieren
van de huid en de hypofyse;
Mesoderm: collageen, spierweefsel, de nieren, de milt, het bloed, bot en het
kraakbeen;
Endoderm: de eicellen, het sperma, de lever, de longen, de blaas, de schildklier,
de darmen en de thymus.
Ook ontstaat uit het ectoderme kiemweefsel de neurale lijst. De neurale lijst (neural
crest) kan worden gezien als de vierde kiemlaag. De neurale lijst is een groep mobiele
neuro-ectodermale cellen die los komen van de ontwikkelende neurale buis (neural
tube).
1.Epitheelweefsel
Epitheelweefsel (epithelial tissue) bestaat uit aaneengesloten cellen waartussen amper
extracellulaire matrix aanwezig is. Ze zijn verbonden door speciale verbindingen die
zorgen voor aanhechting en intercellulaire communicatie. Epitheelweefsel beschermt het
onderliggende weefsel en reguleert de absorptie en secretie van verschillende stoffen. Dit
gebeurt door middel van speciale verbindingen. Zonula occludens (tight junctions) zijn
gebieden in het epitheelweefsel, waarbij de laterale membranen van aangrenzende
epitheelcellen zijn gefuseerd. Hierdoor ontstaat er adhesie en is er controle over de
intercellulaire uitwisseling van moleculen in de nauwe spleet tussen de cellen. De zonula
adherens (adhering junction) is een verbinding die ervoor zorgt dat de epitheelcellen aan
elkaar blijven plakken. Bij deze verbinding zijn de membranen niet gefuseerd, maar is er
een ruimte tussen de cellen die gevuld is met verbindingseiwitten. Dit zorgt voor een
sterke verbinding tussen de cellen. Epitheelcellen kunnen met elkaar communiceren
doormiddel van gap junctions. Dit zijn kleine poriën/kanalen die zorgen voor intercellulair
transport.
Facebook.com/SlimStuderen
22
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Epitheelweefsel wordt vaak ondersteund door bindweefsel (connective tissue). Het
epitheelweefsel bevat geen bloedvaten, maar wordt doormiddel van diffusie via het
bindweefsel van voedingsstoffen voorzien. Epitheelweefsel wordt continue vernieuwd
door de aanvoer van nieuwe cellen van onderaf. Het is daarom dynamisch.
Er zijn verschillende vormen van epitheelweefsel. Simpel epitheelweefsel bestaat maar
uit een enkele laag cellen, hierbij horen plaveiselcelepitheel (squamous epithelium),
kubisch epitheel (cuboidal epithelium), meerrijig epitheel en cilindrisch epitheel
(columnar epithelium). Pseudo-gestratificeerd epitheelweefsel bestaat ook maar uit een
enkele cellaag, maar doordat de kernen op verschillende hoogten liggen lijkt het uit
meerdere cellagen te bestaan. Gestratificeerd epitheelweefsel (stratified epithelium)
bevat twee of meer cellagen. Ook kubisch en cilindrisch epitheel kunnen meerdere
cellagen bevatten. Overgangsepitheel (transitional epithelium) zit tussen de overgang
van gestratificeerd plaveiselcelepitheel en gestratificieerd cilinidrisch epitheelweefsel.
Eenlagig epitheel:
Plaveiselepitheel: bloedvaten, longblaasjes;
Kubisch epitheel: buitenste oppervlak ovaria;
Meerrijig epitheel: luchtpijp;
Cilindrisch epitheel: darmen.
Meerlagig epitheel:
Pseudo-gestratificeerd epitheel: bronchiën;
Gestratificeerd epitheel: epidermis;
Kubisch epitheel: zweetklieren;
Cilindrisch epitheel: conjuctiva oog;
Overgangsepitheel: blaas.
Bovenstaande vormen van epitheel behoren allemaal tot het bedekkende epitheel.
Bedekkend epitheel zijn één of meer lagen van aaneengesloten cellen die het
lichaamsoppervlak of inwendige holten bekleden. Ook is er epitheel dat is gespecialiseerd
in de productie van vloeibaar secreet, dat een functie uitoefent buiten de cel. Exocriene
klieren zijn verbonden met afvoerbuizen en hebben op deze manier een verbinding met
het oppervlakte epitheel. Deze klieren geven hun secreet af aan het lumen. Endocriene
klieren liggen los in het bindweefsel en beschikken hierdoor niet over afvoerbuizen. Ze
lozen hun secreet direct in de bloedbaan.
2.Bindweefsel
Bindweefsel(connective tissue) heeft rondom de cellen een extracellulaire matrix van
vezels en een gel-achtige basis. Bindweefsel ondersteunt en verbindt verschillende
weefsels. Bot, kraakbeen (cartilage), pezen (tendons), gewrichtsbanden (ligaments), de
omhulsels van de organen en vet zijn gespecialiseerde vormen van bindweefsel.
Bindweefsel bestaat uit fibroblastcellen, collageen, elastine, elastische vezels en een
matrix van macromoleculen, zoals suikers, eiwitten en water. Afhankelijk van de
compositie van het bindweefsel kan het zorgen voor kracht of elasticiteit. Tevens kan
bindweefsel de diffusie van voedings- of afvalstoffen tussen het bloed en het
aangrenzende weefsel ondersteunen. Andere functies van het bindweefsel zijn de
bescherming tegen micro-organismen, een steunfunctie en een herstelfunctie na
beschadiging van weefsel.
Er zijn
verschillende typen bindweefselcellen;
fibroblastcellen;
vetcellen (adipose cells);
andere ondersteunende cellen, zoals chondroblasten (produceren kraakbeen),
osteoblasten (ondersteunen het bot), myoblasten (vormen spiercellen);
bewegende cellen uit het bloed (bijvoorbeeld leukocyten);
Facebook.com/SlimStuderen
23
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
mesenchymale cellen, deze cellen kunnen zich differentiëren tijdens de
embryonale fase in alle typen definitieve bindweefselcellen, dit kan ook later ook
nog regeneratie.
Bindweefsel is onder te verdelen in twee typen: het losmazige bindweefsel (loose
connective tissue) en het straffe bindweefsel (dense connective tissue). Het losmazige
bindweefsel bevat weinig cellen, maar veel extracellulair materiaal. Het losmazige
bindweefsel zorgt voor de ondersteuning van organen. Vetweefsel is een voorbeeld van
losmazig vetweefsel. Straf bindweefsel is veel dichter en steviger dan losmazig
bindweefsel. Onregelmatig straf bindweefsel bevat collageen wat in alle richtingen voor
stevigheid zorgt. Regelmatig straf bindweefsel bevat collageenbundels die parallel in
regelmatige richtingen voor nog meer stevigheid zorgen. Een voorbeeld hiervan zijn
pezen.
3.Spierweefsel
Spierweefsel is op te delen in drie verschillende soorten:
Skeletspierweefsel: dwarsgestreepte, bevat veel kernen en evenwijdige
spiercellen. Ze zorgen voor een snelle en krachtige contractie die afhankelijk is
van de wil.
Hartspierweefsel: eenkernige, dwarsgestreepte spiercellen. Deze spiercellen
liggen niet zoals skeletspierweefsel evenwijdig, maar hebben vertakkingen. De
contractie is ritmisch en onder invloed van het autonome zenuwstelsel.
Gladspierweefsel: geen dwarsgestreepte spiercellen. De contractie is langzaam en
onder invloed van het autonome zenuwstelsel.
4.Zenuwweefsel
Het zenuwweefsel vormt het brein, het ruggenmerg, de perifere zenuwen, de ganglia en
speciale receptoren.
Het zenuwstelsel is verdeeld in het centrale zenuwstelsel (central nervous system,
CNS) en het perifere zenuwstelsel(peripheral nervous system, PNS). Het centrale
zenuwstelsel bestaat uit de hersenen en het ruggenmerg(spinal cord). Het perifere
zenuwstelsel bestaat uit alle zenuwen die buiten het centrale zenuwstelsel liggen, zoals
de 12 paren craniale zenuwen (cranial nerves), de 31 paren spinale zenuwen (spinal
nerves) en het perifere autonome zenuwstelsel (autonomic nervous system, ANS). Het
perifere zenuwstel wordt naar functie verdeeld in het autonome en het somatische
zenuwstelsel. Het autonome zenuwstelsel is opgedeeld in het parasympatische
zenuwstelsel en het sympathische zenuwstelsel. Het parasympatische zenuwstelsel is
actief tijdens rust. Het sympathische zenuwstelsel is actief tijdens stressvolle situaties en
fysieke activiteiten. Het somatische zenuwstelsel heeft controle over de interacties met
de buitenwereld en staat, in tegenstelling tot het autonome zenuwstelsel, onder invloed
van de wil.
Het zenuwweefsel van het perifere zenuwstelsel bestaat uit de afferente, sensorische
zenuwen (sensory nerves) en de efferente, motorische zenuwen (motor nerves).
Sensorische zenuwen brengen impulsen over van een weefsel of een orgaan naar het
centrale zenuwstelsel. Motorische zenuwen van het somatische zenuwstelsel geleiden
impulsen van het centrale zenuwstelsel naar de spieren.
! Aanvoerend = afferent
! Afvoeren = efferent
Neuronen
Zenuwcellen worden in de geneeskunde neuronen genoemd. Ze bestaan uit een
cellichaam (perikaryon), met een kern omringd door cytoplasma, een axon en vele
dendrieten. Er bestaan drie primaire typen neuronen: de multipolaire neuronen, de
bipolaire neuronen en de pseudo-unipolaire neuronen. Multipolaire neuronen hebben een
Facebook.com/SlimStuderen
24
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
axon en veel dendrieten. De bipolaire neuronen hebben één axon en één dendriet. De
pseudo-unipolaire neuronen hebben een axon die vertakt in de vorm van een T. De ene
vertakking gaat naar het ruggenmerg, de andere naar het perifere weefsel.
De dendrieten geleiden elektrische signalen, naar het cellichaam. De axonen zijn de
verwerkers van de cel en geleiden impulsen weg van het cellichaam. Dit weggeleiden van
een impuls kan bijvoorbeeld naar een ander neuron, een spiercel of een klier.
Synaps
Tussen het axon van het ene cellichaam en de dendriet van het andere cellichaam zit een
synaps. In deze spleet vindt signaaloverdracht plaats door middel van neurotransmitters.
Dit zijn stofjes die vrijkomen uit het axon(presynaptisch). Ze binden vervolgens op
specifieke structuren van de te stimuleren dendriet (postsynaptisch). Neurotransmitters
liggen in blaasjes opgeslagen aan de onderkant van het axon. Door een elektrische
stimulus die door het axon loopt, komt de neurotransmitter vrij door middel van
exocytose.
Motorische neuronen stimuleren hun doelcel, weefsel of orgaan. Sensorische neuronen
ontvangen prikkels vanuit de weefsels. Schakelneuronen (interneurons/association
neurons) verbinden motorische en sensorische neuronen in het centrale zenuwstelsel.
Gliacellen
Gliacellen ondersteunen de neuronen en dragen bij aan de homeostase van het centrale
zenuwstelsel, maar kunnen geen elektrische prikkels overdragen. Ze zorgen onder
andere voor de versteviging van de hersenen, de productie van myeline en het in stand
houden van de bloed-hersen barrière. Cellen die behoren tot de gliacellen zijn de
astrocyten, de oligodendrocyten, microglia en ependymale cellen.
Astrocyten (astrocytes) helpen met het reguleren van zenuwimpulsen door het
inactiveren van neurotransmitters en helpen met het creëren van een dichte laag voor de
bloed-hersen barrière.
Microglia zijn gespecialiseerd in het fagocyteren van afvalstoffen en beschadigde cellen
in het centrale zenuwstelsel. Ependymale cellen omlijnen de ventrikels van het brein en
het centrale kanaal van het ruggenmerg.
Oligodendrocyten omringen verschillende axonen in het centrale zenuwstelsel en
zorgen zo voor een myeline schede. Schwanncellen wikkelen zich om axonen in het
perifere zenuwstelsel om zo ze te omringen met Schwanncelmembranen, dit wordt de
myeline schede (myelin sheath) genoemd. Er ontstaan gaten tussen de Schwanncellen,
deze worden insnoeringen/knopen van Ranvier (node of Ranvier) genoemd. Deze
vorming van myeline schede gebeurt niet bij alle zenuwen, Schwanncellen kunnen ook
niet-gemyelineerde cellen omringen. Door de vorming van een myeline schede en de
knopen van Ranvier kunnen de potentialen van knoop naar knoop “springen”. Hierdoor
verloopt de overdracht van het signaal sneller.
LET OP: Schwanncellen zorgen voor een myeline schede in het perifere zenuwstelsel,
terwijl oligodendrocyten zorgen voor een myeline schede in het centrale zenuwstelsel.
Eén oligodendrocyt kan tientallen axonen omringen, dit kan de Schwanncel niet.
Schwanncellen omringen elk een klein stuk van één axon.
Aangeboren huidziekten
Epidermolysis bullosa (EB)
Epidermolysis bullosa ontstaat door erfelijke afwijkingen in eiwitten die verantwoordelijk
zijn voor de verbinding tussen de opperhuid en de lederhuid. Het klinische beeld
Facebook.com/SlimStuderen
25
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
behorende bij deze ziekte is de vorming van blaren, vaak verschijnen de blaren al kort na
de geboorte. Er zijn drie varianten:
Epidermolysis bullosa simplex veroorzaakt oppervlakkige blaren door een mutatie
in de eiwitten van het cytoskelet van de basale laag van de epidermis. De
mutatie zit in de chromosomen 12q en 17q, deze coderen voor de eiwitten
keratine 5 en 14. Dit is een milde vorm van EB en vaak zijn er alleen blaren te
zien op de handen en de voeten.
Epidermolysis bullosa dystrophica wordt gekenmerkt door diepere blaren,
littekenvorming en milia formatie (cystes van keratine). Dit wordt veroorzaakt
door een mutatie in het COL-7A1 gen, deze zit op het chromosoom 3p21.1. Deze
mutatie zorgt voor een vermindering van het collageen in de ankerfibrillen
(anchoring fibrils). Ankerfibrillen zorgen ervoor dat het basale lamina van de
epidermis aan de onderliggende dermis zit. Ook de nagels, mucosa en het
strottenhoofd (larynx) worden aangetast. De autosomale dominante variant is
milder dan de autosomale recessieve variant. De recessieve variant zorgt voor
een ernstige ziekte met pijnlijke littekens, gewrichtscontracturen, dysphagia
(moeite met slikken) en dat de tenen en vingers aan elkaar groeien. De
levensverwachting verminderd door deze ziekte erg. Door het herhaaldelijk
ontstaan van littekens is de kans op het ontwikkelen van een
plaveiselcelcarcinoom(squamous cell carcinomas) sterk verhoogd.
Epidermolysis bullosa junctionalis is de meest ernstige vorm van deze ziekte. Het
wordt gekenmerkt door een splijting in de lamina lucida van de basale
membraan. Dit wordt veroorzaakt door mutaties in genen die coderen voor
verschillende eiwitten, voornamelijk in laminine-332. Het is al zichtbaar na de
geboorte door veel grote blaren en gebieden zonder huid. Er zijn ook vaak
afwijkingen in de tanden en de nagels. Er bestaat een dodelijke en een nietdodelijke vorm van EB-junctionalis, maar ze ontstaan beide door een autosomale
recessieve erfelijke afwijking.
De behandeling van EB vindt plaats in een gespecialiseerd centrum. De exacte diagnose
kan worden gesteld door het analyseren van de blaren, maar het wordt echt vastgesteld
door genetische testen. Prenatale diagnose en pre-implantatie is mogelijk voor de
ernstige vormen van EB. Voor epidermolysis bullosa dystrophica wordt stamcel
transplantatie gebruikt. Nieuwe mogelijkheden die worden onderzocht zijn gentherapie
en beenmerg transplantatie.
Facebook.com/SlimStuderen
26
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
E. FYSIOLOGIE
Zenuwfysiologie
Globale functie van het centrale zenuwstelsel
In het lichaam vinden allerlei razendsnelle processen plaats om signalen door te geven.
De signalen worden door middel van zenuwen vervoerd naar het centraal zenuwstel. Het
doorgeven van deze signalen gebeurt op een bijzondere manier. Belangrijke begrippen in
dit aspect zijn het rustmembraanpotentiaal en het actiepotentiaal.
Rustmembraan potentiaal
Er bestaat altijd een spanning in neuronen, zelfs wanneer er geen impulsen zijn. Dit
wordt de rustpotentiaal genoemd. Deze rustpotentiaal ontstaat door een oneerlijke
verdeling van Na+- en K+- ionen. Er zitten namelijk meer K+-ionen en minder Na+- ionen
binnen in de cel, dan buiten de cel. De rustpotentiaal is ongeveer -90 mV. Om het rust
potentiaal in stand te houden beschikt de cel over ‘lek’ kanalen. Deze laten veel liever
K+-ionen door dan Na+-ionen. De ‘lek’ kanalen zijn passief en werken op de
concentratiegradiënt. Daarnaast beschikt de cel over een actieve Na +/K+-pomp. Onder
invloed van ATP wordt hierdoor twee K+ naar binnen gepompt en drie Na+ naar buiten.
Dit veroorzaakt het negatieve potentiaal aan de binnenzijde van het membraan.
Actiepotentiaal
Als gevolg van een prikkel kunnen prikkelbare cellen, zoals bijvoorbeeld spier- en
zenuwcellen, de ionenpermeabiliteit van hun celmembraan veranderen. Stimulatie van
een neuron zorgt er voor dat plaatselijk de membraanpermeabiliteit voor natriumionen
wordt verhoogd: het actiepotentiaal. Er ontstaat een actiepotentiaal wanneer de
potentiaal een kritische waarde bereikt. De drempelwaarde is normaal 8-12 mV lager dan
de rustmembraanpotentiaal.
Belangrijke kanalen voor de generalisatie van het actiepotentiaal zijn de voltage-gated
kanalen. Hierbij is het voltage-gated natrium kanaal verantwoordelijk voor de
depolarisatie en het voltage-gated kalium kanaal voor de repolarisatie. De Na+/K+pomp is gedurende het actiepotentiaal verantwoordelijk voor het herstellen van het
potentiaal.
Wanneer een actiepotentiaal start springen de voltage-gated natrium kanalen open. Na+
gaat hierdoor de cel binnen en veroorzaakt een depolarisatie van het
membraanpotentiaal. Na slechts enkele milliseconden sluiten de voltage-gated natrium
kanalen weer. De voltage-gated kalium kanalen open iets langzamer en blijven ook
langer open, repolarisatie treedt op. Wanneer het potentiaal weer negatief wordt sluiten
de voltage-gated kalium kanalen. Wanneer dit gebeurt met een kleine vertraging wordt
gesproken over een hyperpolarisatie.
De refractaire periode is de periode waarin de voltage-gated natrium kanalen niet
kunnen openen. Tijdens deze periode kan de cel geen nieuw actiepotentiaal verwerken.
De refractaire periode stopt wanneer het membraanpotentiaal zijn standaardwaarde weer
bereikt.
Als een zenuwvezel plaatselijk depolariseert, wordt er een stroom gestuurd van het
gedepolariseerde punt naar de aangrenzende microregio. Dit komt doordat er bij het
gedepolariseerde punt een ladingsomkering heeft plaatsgevonden en er is dus een
positieve lading aan de binnenzijde van het membraan. Het is echter zo, dat het
membraan verderop nog negatief is. Hierdoor stromen positieve ladingen van de
gedepolariseerde positieve regio naar de negatieve regio stromen. Het proces herhaalt
zich nu constant en het actiepotentiaal loopt over de gehele lengte van de zenuwvezel.
Facebook.com/SlimStuderen
27
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Myeline
Een speciaal kenmerk bij zenuwen om een signaal zo snel mogelijk door te geven is de
isolatie van de zenuw. Lange zenuwen zijn gemyeliniseerd, korte zenuwen
ongemyeliniseerd. Myeline is verantwoordelijk voor de isolatie bij de voortgeleiding van
actiepotentialen. De geleiding over een axon kan hierdoor veel sneller verlopen. Het
actiepotentiaal kan door het myeline namelijk tussen de knopen van Ranvier (node of
Ranvier) “springen” en zich zo sneller over het axon verplaatsen.
Het somatische zenuwstelsel
Het somatische zenuwstelsel, ook wel het vrijwillige zenuwstelsel genoemd, regelt alle
interacties met de buitenwereld. De sensorische neuronen brengen de boodschappen
over vanuit de organen die het waarnemen (bijvoorbeeld de huid). De motorische
sensoren activeren de skeletspieren. Het somatische zenuwstelsel wordt gevormd uit de
somatische delen van het centraal en perifere zenuwstelsel en zorgt voor algemene
zintuigelijke en motorische innervatie voor alle delen van het lichaam, behalve voor de
ingewanden in de lichamelijke holten, de gladde spieren en de klieren.
Het autonome (viscerale) zenuwstelsel is het deel van het perifere zenuwstelsel dat voor
de reacties zorgt die onbewust plaatsvinden, zoals het ademhalen en vele functies van
organen zoals de lever en nieren.
Spierweefsel
Iedere spiervezel bevat honderden tot duizenden myofibrillen (myofibrils). Elk myofibril
bestaat uit een afwisseling van ongeveer 1500 myosinefilamenten en 3000
actinefilamenten. A-banden bevatten dikke myosinefilamenten die aan de uiteinden
overlappen met dunne actinefilamenten. In het midden is geen sprake van een
overlapping met myosine filamenten, dit wordt de H-band genoemd. De I-band bevat
myosine en actine die elkaar niet overlappen. Dit is de reden waarom deze band op
weergaves vaak lichter van kleur is. In het midden zit de Z-band, waar de
actinefilamenten aanhechten. Het deel dat zich tussen de Z-banden bevindt wordt het
sarcomeer genoemd.
De zij-aan-zij ligging van actine en myosine is moeilijk te verkrijgen. Het wordt op de
juiste plaats gehouden door het eiwit genaamd titin. Het is draadvormig, en hierdoor is
het eiwit erg elastisch. Het eiwit houdt het myosine en actine op zijn plaats, waardoor
het samentrekkende systeem van het sarcomeer kan werken. Titin is verbonden aan de
Z-band, en verandert van lengte bij contractie en ontspanning.
Mitochondriën liggen dichtbij de myofibrillen voor energietoevoer. De ruimten tussen de
myofibrillen wordt gevuld met een vloeistof; het sarcoplasma (sarcoplasm).
De initiatie en uitvoering van spiercontractie vindt plaats in de volgende stappen:
Er wordt een actiepotentiaal afgegeven door een motorische neuron, dat uitmondt
bij een spiervezel.
De zenuw scheidt de neurotransmitter acetylcholine uit bij elk zenuwuiteinde.
Acetylcholine zorgt er voor dat in het spiervezelmembraan ion-kanalen open gaan.
Deze opening van de kanalen zorgt er voor dat er grote hoeveelheden
natriumionen het membraan in kunnen gaan. Hierdoor treedt depolarisatie op van
het sarcolemma, het membraan van de spiervezel. Dit leidt tot opening van
spanningsgevoelige natriumkanalen.
De depolarisatie wordt via T-tubuli doorgegeven aan de myofibrillen.
Depolarisatie in de myofibrillen leidt tot activatie van het sarcoplasmatisch
reticulum om grote hoeveelheden calcium vrij te geven, die hierin zijn gemaakt.
De calciumionen binden aan het troponine complex. Deze veranderd hierdoor van
vorm en er komen bindingsplaatsen vrij. De koppen van het mysone filament
kunnen op de vrijgekomen plaatsen binden.
Facebook.com/SlimStuderen
28
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
ATP komt vrij wanneer de koppen gebonden zijn. De koppen kunnen hierdoor
buigen, waardoor de actine en myosine filamenten langs elkaar worden
getrokken. Dit is de het proces van samentrekken.
De samentrekking wordt beëindigd doordat de calciumionen worden
teruggepompt naar het sarcoplasmatisch reticulum.
Een myosinefilament bestaat uit zes polypeptideketens; twee zware en vier lichte ketens.
De twee zware ketens vormen een dubbele helix; dit wordt de staart van het
myosinemolecuul genoemd. Een einde van een van deze ketens is bilateraal gevouwen in
een bolvormige polypeptidestructuur; dit wordt de myosinekop (myosin head)
genoemd. Er zijn dus twee myosinebolletjes aan het eind van een dubbele helix. De vier
lichte ketens maken deel uit van de bolletjes; twee op elke myosinekop.
In toestand van rust overlappen de actine- en myosinefilamenten elkaar gedeeltelijk. Bij
contractie verplaatsen deze filamenten ten opzichte van elkaar. De myosinefilamenten
schuiven nu tussen de actinefilamenten; dus gedurende de contractie schuift het hele
pakket van myosine- en actine filamenten van een sarcomeer in elkaar. Dit gebeurt
allemaal in de A-band. Wanneer een spier in rust is, wordt de bindingsplaats tussen
myosine en actine geblokkeerd door het troponine-tropomyosinecomplex op het actine
filament. Hierdoor wordt voorkomen dat er contractie plaatsvindt.
Er zijn drie soorten spierweefsel. In onderstaande tabel staan de soorten die er worden
onderscheiden.
Skeletspierweefsel
Lange en cilindervormige
veelkernige reuscellen die
parallel gerangschikt zijn
Myoblasten (stamcellen)
vormen door deling en fusie
een veelkernige spiercel
Hartspierweefsel
Eenkernig,
dwarsgestreept en
vertakt
Vertakken en
hechten zich met de
uiteinden van de
vertakkingen aan
elkaar
Streping
Dwarse streping door
evenwijdige rangschikking
Dwarse streping
Ligging
kern
Regeneratie
Direct onder het
sarcolemma gelegen
Door satellietcellen (een
eenkernige stamcel die bij
deling met elkaar of een
andere spiercel kan fuseren)
Centraal in de cel
Uiterlijke
kenmerken
Vorming
Niet mogelijk
Gladde spiercellen
Spoelvormig
Spiercellen zijn aan
elkaar gekoppeld
door middel van
dense bodies. Per
spiercel maar een
celkern
Geen dwarse
streping. Actine,
myosine en dense
bodies zijn wel
aanwezig
In het midden van
de cel
Door celdeling
mogelijk (ook
satellietcellen
aanwezig)
Door twee verschillende stoffen kan de werking van de motorische eindplaat beïnvloed
worden.
Cholinesteraseremmers: deze stof vertraagt de afbraak van acetylcholine in de
synapsspleet en verlengt daardoor de werking van de transmitter. Het is hierdoor
mogelijk dat een spasme ontstaat in de spier.
Curare: deze stof blokkeert de binding van acetyolcholine aan het postsynaptisch
membraan. Het gevolg hiervan is dat niet genoeg poorten openen om het
actiepotentiaal te laten ontstaan. Er vindt geen contractie plaats.
Facebook.com/SlimStuderen
29
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Een isometrische contractie(isometric contraction) is een contractie waarbij er kracht
wordt geleverd zonder dat de lengte van de spier verandert. Een isotone
contractie(isotonic contraction) is een contractie met een gelijke spierspanning.
De kracht die geleverd wordt bij een contractie hangt af van meerdere factoren.
Essentieel om te weten is of de spier wordt gestimuleerd in kleine schokjes of constant
wordt gestimuleerd. Dit noemt men een twitch of een tetanus.
Een twitch is de tijd tussen het motorisch eindplaat potentiaal, de contractie en de
ontspanning (relaxatie) van een spiervezel. Tetanus veroorzaakt een constante contractie
zonder relaxatie van de spiervezel.
Facebook.com/SlimStuderen
30
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
F. PATHOLOGIE
Cellulaire respons op stress en toxines
Cellen nemen toe in grote tijdens de G1 en G2 fase, door de toename van cytoplasma en
organellen en genetisch materiaal voor de replicatie tijdens de S-fase. Door de celdeling
nemen de cellen toe in aantal.
Cel differentiatie is het proces waarbij niet gespecialiseerde cellen steeds
gespecialiseerder worden. Dit gebeurt tijdens de gehele ontwikkeling van organismen,
van een simpele zygoot tot een gespecialiseerd organisme. Deze veranderingen worden
veroorzaakt door gecontroleerde aanpassingen in de expressie van genen.
Toch heeft een gespecialiseerde cel, die beperkt is tot een aantal functies en een
bepaalde structuur door zijn metabolisme, differentiatie en specialisatie, de mogelijkheid
zich aan te passen en tegelijk de homeostase te bewaren. Dit gebeurt door middel van
adaptaties. Adaptaties zijn veranderingen in de grote, het aantal, het fenotype,
metabolische activiteit of functie van cellen in reactie op veranderingen in hun omgeving.
Dit kan door in grote, activiteit en aantal toe te nemen, in activiteit af te nemen of door
een verandering in het fenotype van de cellen. Dit kan gebeuren in een aantal vormen.
Hypertrofie(hypertrophy)
Hypertrofie is een toename in de grote van cellen, waardoor het orgaan ook in grote
toeneemt. In reactie op mechanische stress of ander stimuli worden er groeifactoren
geproduceerd die zorgen voor een toename van de productie van cellulaire eiwitten.
Hypertrofie kan pathologisch zijn, bijvoorbeeld cardiale hypertrofie in de hartspier door
een te hoge bloeddruk. Het kan echter ook fysiologisch zijn, bijvoorbeeld de groei van de
baarmoeder tijdens zwangerschap. Hypertrofie vindt plaats in weefsels waar geen
celdeling kan plaatsvinden.
Hyperplasie(hyperplasia)
Hyperplasie is de toename van het aantal cellen in een orgaan of weefsel in reactie op
een stimulus. Hyperplasie kan dus alleen voorkomen bij cellen die in staat zijn zich te
delen. Fysiologische hyperplasie wordt veroorzaakt door hormonen of andere
groeifactoren in verschillende omstandigheden. Bijvoorbeeld wanneer er een toename
nodig is in de capaciteit van hormoongevoelige organen of wanneer er compensatie nodig
is na schade. Hierbij hoort bijvoorbeeld ook de groei van borsten tijdens de puberteit. De
meeste vormen van pathologische hyperplasie worden veroorzaakt door een te grote of
ongepaste activiteit van hormonen of groeifactoren. Hypertrofie gaat vaak samen met
hyperplasie. Hyperplasie vindt plaats in weefsels die in staat zijn zich te delen of
stamcellen bevatten.
Atrofie(atrophy)
Atrofie is een afname van de grote van een orgaan of weefsel door een afname van het
aantal cellen en de grote van cellen. Fysiologische atrofie is normaal en vindt plaats
tijdens de foetale ontwikkeling. De meest voorkomende oorzaken van pathologische
atrofie zijn:
verminderde werklast;
schade aan zenuwen;
ischemie (vermindering van bloedtoevoer);
onvoldoende voeding;
vermindering van de endocriene stimulatie (hormoonstimulatie);
druk (druk op weefsels door bijv. een tumor).
Atrofie wordt veroorzaakt door een afname van de synthese van eiwitten en andere
cellulaire bouwstenen en een toename in de afbraak van organellen.
Facebook.com/SlimStuderen
31
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Metaplasie (metaplasia)
Metaplasie is een omkeerbare verandering waarbij een gedifferentieerd celtype (epitheel
of mesenchymaal) wordt vervangen door een ander celtype. De meest voorkomende
epitheel metaplasie is cilindrisch naar plaveiselcelepitheel. Dit gebeurt bijvoorbeeld in de
luchtwegen bij rokers.
Metaplasie wordt niet veroorzaakt door een verandering in het fenotype van een
gedifferentieerde cel. In het geval van metaplasie wordt de cel opnieuw geprogrammeerd
vanuit stamcellen die in normaal weefsel zitten of van ongedifferentieerde mesenchymale
cellen uit bindweefsel. Het gevolg hiervan kan zijn dat het weefsel niet goed meer kan
functioneren of dat er een grotere kans is op kwaadaardige transformaties (kanker).
Celschade
Cel zwelling is de eerste manifestatie van bijna alle vormen van schade aan cellen. Dit is
het beste te zien aan een geheel orgaan door bleekheid (pallor), een toegenomen
turgor(water gaat via osmose de cel in) en een toename van het gewicht van het orgaan.
Door middel van microscopie kunnen kleine vacuolen worden gezien in het cytoplasma,
dit zijn opgezwollen en afgeknelde stukken van het ER. Deze zwelling van cellen is
omkeerbaar. In cellen die te maken hebben met vetmetabolisme wordt vaak steatose
(fatty change) gezien, dit kan worden gezien door de verschijning van lipide vacuolen in
het cytoplasma.
De veranderingen die te zien zijn met een elektronenmicroscoop (ultrastructureel) zijn:
blaarvorming, afstomping en afname van microvilli in het plasmamembraan;
zwelling en verschijning van amorphous densities in mitochondriën;
vergroting van het ER en het los komen van polysomen;
DNA schade en andere schade in de kern.
Celdood
Wanneer een cel beschadigd wordt en wanneer dit te lang duurt, kan de cel zich niet
meer herstellen. Hierdoor zal de cel dood gaan. Door deze schade functioneert de
celwand niet goed meer en zullen er organellen en cytoplasmavloeistoffen naar buiten
stromen. Dit veroorzaakt een ontstekingsreactie in het omliggende weefsel. De enzymen
die de cel verteren komen uit de lysosomen van de cel of uit de leukocyten die onderdeel
zijn van de ontstekingsreactie. Dit proces heet necrose. Necrose leidt tot schade aan de
omliggende weefsels.
Apoptose is gecontroleerde celdood. Cellen die bestemd zijn om dood te gaan, activeren
enzymen binnen in de cel. Hierdoor worden het DNA en de eiwitten van de cel
afgebroken. De resten van de cel worden in vacuolen verpakt(apoptotic bodies) en
afgesnoerd door de celwand. Deze vacuolen worden gefagocyteerd door fagocyten.
Doordat er geen onderdelen van de cel los in het weefsel terecht komen, ontstaat er
geen ontstekingsreactie. Apoptose leidt dus niet tot schade aan de omringende weefsels.
Apoptose is essentieel in de ontwikkeling van een embryo, door apoptose verdwijnen de
ongewenste weefsels. Apoptose komt ook voor na een immuunreactie. Wanneer
leukocyten en neutrofielen hun taak hebben volbracht ondergaan ze apoptose.
Autofagie is een proces waarbij een cel zijn eigen onderdelen opeet. Wanneer er niet
genoeg voedingstoffen zijn, kan een cel een dubbel membraan (autophagosome) om
materialen uit het cytoplasma maken. De autofagosoom fuseert met een lysosoom en
breekt op deze manier zijn inhoud af. Autofagie is een reactie op een tekort aan
voedingsstoffen en dus een soort overlevingsmechanisme.
Weefselvernieuwing, -regeneratie en –herstel
Cel proliferatie is fundamenteel voor de ontwikkeling, behoud van homeostase en het
vervangen van dode of beschadigde cellen. Hiervoor is een goede DNA-replicatie en een
Facebook.com/SlimStuderen
32
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
goede synthese van organellen belangrijk, gevolgd door een goede verdeling tussen de
dochtercellen.
Groeifactoren en signaalmechanismen bij celgroei
Groeifactoren stimuleren de activiteit van genen die nodig zijn voor de groei van cellen
en celdeling. Groeifactoren kunnen aan specifieke receptoren binden die invloed hebben
op de expressie van genen die ervoor zorgen dat:
cellen de celcyclus binnengaan;
blokkades van de celcyclus weg worden gehaald;
er geen apoptose plaatsvindt;
de biosynthese van celonderdelen wordt gestimuleerd.
Signaalmechanismen zorgen ervoor dat een cel kan communiceren met zijn omgeving.
Dit is erg belangrijk voor de regulatie van de celgroei. Sommige signalen stimuleren een
cel om zich te differentiëren, andere stimuleren proliferatie en weer andere signalen
zorgen ervoor dat de cel een gespecialiseerde functie gaat uitvoeren. De communicatie
tussen cellen is belangrijk in embryo’s, maar ook wanneer er schade optreedt in een
weefsel. Wanneer cellen deze communicatie niet hebben, leidt dit tot ongereguleerde
groei (kanker) of tot een soort shock, waarbij de cel niet reageert op zijn omgeving.
Stamcellen
Embryonale stamcellen zijn de minst gedifferentieerde cellen. Ze zitten in de
blastocyste en kunnen zich nog differentiëren tot elke cel in het lichaam, inclusief de
placenta. Door deze mogelijkheid worden ze totipotent genoemd.
Volwassen somatische stamcellen(adult somatic stem cells) zijn stamcellen die niet
meer elke cel kunnen worden. Volwassen somatische stamcellen in een bepaald weefsel
kunnen vaak alleen cellen produceren die normaal zijn voor dat type weefsel. Volwassen
somatische stamcellen worden vaak beschermd in een micromilieu(stem cell niches).
Deze omgeving zorgt ervoor dat de stamcel rustig blijft tot het moment dat het nodig is
dat de cel zich gaat differentiëren.
iPS cellen(induced pluripotent stem cells) zijn gespecialiseerde volwassen cellen die via
kunstmatige herprogrammering weer pluripotent worden, dit houdt in dat ze kunnen
uitgroeien tot alle typen lichaamscellen. De herprogrammering gebeurt door aan het DNA
van de cel regulatorgenen toe te voegen en die tot expressie te brengen. Deze
regulatorgenen zijn in gedifferentieerde cellen niet actief, maar wel in embryonale
stamcellen.
Weefsel- en orgaanregeneratie
Er zijn verschillende mechanismen betrokken bij weefsel- en orgaanregeneratie.
In labiele weefsels zoals het epitheelweefsel van de darmen en de huid, worden cellen
snel vervangen. Dit gebeurt door stamcellen die in het onderliggende basale membraan
zitten. Weefselregeneratie kan in mindere mate ook plaatsvinden in weefsels met een
stabiele cel populatie. De mechanismen hiervoor zijn hypertrofie en hyperplasie.
Permanente weefsels kunnen zich niet meer differentiëren en schade kan niet meer
gerepareerd worden. Neuronen en hartcellen horen bij deze groep.
De lever heeft een stabiele cel populatie, maar kan zich in veel gevallen goed herstellen.
De regeneratie wordt gestimuleerd door cytokines en groeifactoren die geproduceerd
worden in reactie op het verlies aan levermassa en een ontsteking.
Cel proliferatie wordt gecontroleerd door de celcyclus en wordt gestimuleerd door
groeifactoren en interactie van cellen met de extracellulaire matrix.
Wanneer er geen totaal herstel kan plaatsvinden door regeneratie, worden de
beschadigde cellen vervangen door bindweefsel, dit leidt tot littekenweefsel.
Facebook.com/SlimStuderen
33
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
G. RECHT & ETHIEK
De geheimhoudingsplicht
Zowel de maatschappij als de patiënt hebben belang bij de geheimhoudingsplicht.
Het algemeen belang van het beroepsgeheim houdt in dat iedereen zich ongehinderd
naar een arts kan wenden voor hulp en bijstand. Dit draagt bij aan ‘het gelijke recht op
gezondheidszorg’ voor iedereen.
Het individuele belang houdt in dat de patiënt weet dat hij volledig open kan zijn
tegenover zijn arts. Hij weet dat de informatie die hij aan zijn arts verschaft niet zonder
toestemming voor andere doeleinden kan worden gebruikt of aan derden kan worden
gegeven.
De geheimhoudingsplicht omvat alle gegevens, die een arts over de patiënt te weten
komt. Dus ook wanneer niet de patiënt zelf, maar iemand anders, de arts in vertrouwen
heeft genomen over de betreffende patiënt.
Onder de geheimhoudingsplicht vallen de zwijgplicht en het verschoningsrecht. De
zwijgplicht geeft de arts de plicht om te zwijgen over alles wat hem tijdens zijn rol als
dokter over een patiënt bekend wordt. Dit betekent zowel dat hij moet zwijgen over alles
wat hij over een patiënt tijdens een behandeling is te weten gekomen, als dat hij moet
zwijgen over het feit dat de patiënt bij de arts onder behandeling is. Het
verschoningsrecht houdt in dat de arts moet weigeren wanneer hij of zij als getuige
wordt door bijvoorbeeld een rechter of iemand van politie en justitie. Wanneer de arts
een beroep doet op het verschoningsrecht, is het aan de rechter of het beroep zal
worden gehonoreerd. Is dat niet het geval, dan zal de arts alsnog moeten spreken. Het
verschoningsrecht van een arts heeft twee belangrijke consequenties:
Geen aangifteplicht. De arts is op grond van het verschoningsrecht vrijgesteld
van de aangifteplicht. Iedere burger in Nederland heeft deze plicht om aangifte te
doen bij de politie wanneer men weet van een gepleegd ernstig misdrijf.
Geen inbeslagneming dossiers. Bij personen die een verschoningsrecht
hebben, mogen brieven of andere geschriften waarop de geheimhoudingsplicht
rust niet in beslag worden genomen. Een medisch dossier mag dus niet in beslag
worden genomen door politie, openbaar ministerie of rechter commissaris, tenzij
de arts daar toestemming voor geeft. De Hoge Raad heeft echter bepaald dat
inbeslagneming wél mag plaatsvinden in zeer uitzonderlijke omstandigheden,
waarin het belang van de waarheid zo groot is, dat de schending van het
beroepsgeheim gerechtvaardigd is.
De geheimhoudingsplicht kan alleen doorbroken worden als er sprake is van:
Toestemming van de patiënt
Wettelijke plicht tot spreken. Hieronder vallen de Wet op de lijkbezorging en de
Wet publieke gezondheid.
Conflict van plichten. Met het vrijgeven van informatie kan een acuut en direct
gevaar voor de veiligheid of het leven van de patiënt of anderen worden
voorkomen. Bij het maken van een keuze moet de arts zich de volgende vragen
stellen:
- Is het mogelijk toestemming aan de patiënt te vragen?
- Kan de patiënt het probleem zelf oplossen?
- Is er een reëel risico op acuut en direct gevaar?
- Wordt het risico daadwerkelijk weggenomen door politie/justitie te
informeren?
- Is er geen andere weg om de dreiging weg te nemen?
- Is er een redelijke verhouding tussen middel en doel?
Wanneer de arts moet kiezen om de geheimhoudingsplicht wel of niet te verbreken, moet
hij de beginselen van subsidiariteit en proportionaliteit meewegen.
Facebook.com/SlimStuderen
34
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Met subsidiariteit wordt bedoeld dat de arts zich moet afvragen of de veiligheid van het
slachtoffer ook op minder ingrijpende wijze kan worden beschermd dan de politie in te
richten. Wanneer dit het geval is, moet de arts kiezen voor het minder ingrijpende
alternatief. Wanneer direct gevaar alleen kan worden opgeheven door de
geheimhoudingsplicht te verbreken, is dat toegestaan.
Met proportionaliteit wordt bedoeld dat de schade als gevolg van de schending van het
geheimhoudingsplicht in verhouding moet staan tot het voordeel dat de schending met
zich meebrengt. Wanneer deze inschatting lastig is voor de arts, kan hij een
vertrouwelijk overleg plegen met een collega of met de infolijn van de KNMG-artsen.
Algemene principes medische ethiek
Ethiek is een tak van de filosofie die zich bezighoudt met de kritische bezinning over het
juiste handelen. In algemene zin probeert ethiek de criteria vast te stellen om te kunnen
beoordelen of een handeling als goed of fout kan worden gekwalificeerd en om de
motieven en consequenties van deze handeling te kunnen evalueren.
Normen en waarden
Een belangrijk onderdeel van de medische ethiek zijn normen en waarden. Beide
woorden hebben een hele andere, en toch wel met elkaar samenhangende betekenis. Als
er over normen en waarden wordt gesproken, worden de omgangsvormen en principes
bedoeld waarmee men publiekelijk en in sociaal verband voor de dag kan komen.
Normen en waarden geven ons richting en beïnvloeden ons gedrag.
Normen zijn de zaken die als normaal betiteld worden. Het zijn richtlijnen hoe men zich
dient te gedragen. Dit zijn een soort ongeschreven regels van goed fatsoen waar de
meeste mensen zich min of meer aan houdt.
Waarden zijn zaken die waardevol gevonden worden door iemand of een groep mensen,
een samenleving. Vaak is het een enkel woord, zoals eerlijkheid, respect of gelijkheid.
Waar normen gaan over hoe men zich hoort te gedragen, gaan waarden over wat de
achterliggende gewenste motivatie is.
Eigenlijk zijn normen dus afgeleid aan waarden. Eerst is er de waarde, dan volgt de
norm.
Ook belangrijk in de medische ethiek is de rechtvaardiging van de acties van een arts. De
rechtvaardiging is het aantonen van de juistheid of rechtvaardigheid van iets. Met
rechtvaardig zijn, wordt eerlijk, deugdzaam en volgens het recht zijn bedoeld. Zoals aan
de definities te zien, is rechtvaardiging erg subjectief, maar over het algemeen zijn leden
van bevolkingsgroepen het redelijk met elkaar eens wat rechtvaardig is.
Een ander belangrijk ethisch aspect is de maxime. Dit is het principe dat de aanzet geeft
of ten grondslag ligt aan iemands handelingen. Waarom handelt een individu op een
bepaalde manier? Een maxime is een subjectief principe, dat gebaseerd is op de wil van
het individu. In tegenstelling tot de praktische wet die de wil van iedereen bepaalt, is de
maxime het beginsel waarop een individu handelt.
Er zijn verschillende ethische theorieën, zoals:
Utilitarisme: Goed en kwaad is een kwestie van rekenen. Van welke handeling
worden de meeste mensen gelukkig?
Deontologie: Handelingen worden niet gerechtvaardigd door hun gevolgen. De
correctheid van handelingen wordt bepaald door de middelen en niet door goede
gevolgen.
Deugdethiek: Mensen zijn te vormen door opvoeding en onderwijs. Vorm
mensen tot moreel goede verantwoordelijke wezens voor het leiden van een goed
leven en daarvoor is het ontwikkelen van goede eigenschappen, deugden,
noodzakelijk. Voorbeelden van deugden zijn betrouwbaarheid, humor,
verantwoordelijkheid en moed.
Facebook.com/SlimStuderen
35
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
Hippocratische ethiek: Een verantwoordelijke beoefening van de geneeskunde
vraagt van de arts niet alleen deskundigheid, vaardigheid, maar ook waardigheid
en bepaalde morele kwaliteiten. Het ideaalbeeld van een goede
beroepsuitoefening is de medicus gratiosus, de dienstvaardige dochter. De
Hippocratische traditie van medische ethiek beroept zich op deze oorsprong. Ze
gaat terug tot het begin van de geneeskunde zoals wij die nu kennen.
Centrale begrippen medische ethiek
Ethiek vindt plaats op verschillende niveaus, namelijk beschrijvend, normatief en meta;
persoonlijk en professioneel; en individueel.
De beschrijvende ethiek houdt zich bezig met de beschrijving van de heersende
moraal. Hierbij worden zeden en gewoonten, opvattingen over goed en kwaad,
verantwoord en onverantwoord gedrag en geoorloofde en ongeoorloofde handelingen
beschreven.
De normatieve ethiek oordeelt over de moraal. De volgende vraag staat hierbij
centraal: beantwoorden de feitelijke gehanteerde normen en waarden wel aan de
opvattingen over hoe mensen zich behoren te gedragen?
De meta-ethiek reflecteert over wat de achtergronden zijn van onze overtuigingen in
verband met wat we als het goede beschouwen en hoe we onze levens moeten leiden.
Ethiek op persoonlijk niveau gaat om de persoonlijke ethiek van de arts en bij ethiek op
professioneel niveau gaat het om de ethiek die de arts naleeft als arts zijnde. Bij ethiek
op individueel niveau gaat het om de ethiek van een individu.
Vaak komen ethische problemen voor. Enkele typen hiervan zijn:
Conflicterende problemen: er kan niet aan alle verantwoordelijkheden tegelijk
worden gedaan;
Sociaalpsychologische factoren: eigenbelang, gebrek aan bereidheid,
microscopische visie, gehoorzamen aan autoriteiten, toewijding, groepsdenken,
depersonalisatie en onzorgvuldige oordeelsvorming;
Het vele-handenprobleem: gedeelde verantwoordelijkheid, waarbij niet duidelijk is
bij wie de verantwoordelijkheid ligt.
Morele problemen lijken vaak op (technische) ontwerpproblemen. Overeenkomsten zijn:
Beide zijn slecht gestructureerde problemen;
Het ontbreken van en duidelijk criterium waarmee kan worden nagegaan of een
voorgestelde oplossing acceptabel is;
Meerdere oplossingen zijn mogelijk en er is geen definitieve verzameling van alle
mogelijke oplossingen te formuleren;
Het is onmogelijk de probleemruimte volledig te definiëren.
De cyclus waarmee ethische problemen kunnen worden opgelost heet de empirische
cyclus. Deze heeft een aantal kenmerken:
Het probleem oplossen is een trial-and-error proces;
Oplossingen worden veelal niet direct in de werkelijkheid uitgeprobeerd;
Om het probleem op te lossen wordt de cyclus doorgaans vele malen doorlopen.
De cyclus bestaat uit de volgende onderdelen: waarnemen, vermoeden, verwachten,
toetsen en evalueren. Bij het oplossen van morele problemen is het doel de moreel
meest wenselijke handelingsmogelijkheid te bepalen.
Facebook.com/SlimStuderen
36
Overzicht studiestof “De ontwikkeling” blok 1.1.1 2014-2015
BON
10% korting
bij je eerste online aankoop op
http://www.slimstuderen.nl
Gebruik bij het bestellen code GNKRUG2014.
De code is geldig tot en met 14 oktober.
Facebook.com/SlimStuderen
37
TENTAMENS HALEN?!
~ Volg een cursus bij CAPITA SELECTA ~
3,4 of 5 lessen van 3 uur
De beste student-docenten
Kleine klassen
Ruimte voor vragen
Oefenen met (oude) tentamenvragen
Optimaal voorbereid naar jouw tentamen
Inschrijven via www.capitaselecta.nl !
Vragen via [email protected] of 0102142394.
In blok 1 biedt CAPITA SELECTA de volgende
cursussen voor jou aan:
Blok 2.1: Waarnemen en reageren (2 deeltentamens)
Blok 3.1: Oncologie en Bewegingsapparaat (eindtentamen)
Ook zullen er cursussen gegeven worden voor het nieuwe curriculum!