Facts
Series
Banaan
het groene goud
van het Zuiden
Inhoudsopgave
Samenvatting Feiten en cijfers
3
4
Samenvatting
Wereldwijd kent en eet iedereen bananen. Met een jaarlijkse productie van 145 miljoen ton in meer dan
Hoofdstuk 1: Banaan, het groene goud van het Zuiden
5
De voorouders van de moderne banaan
6
Daarom zijn de bananen krom
7
Een gebakken banaan of eentje uit het vuistje?
8
geteeld in Azië, een derde in Latijns-Amerika en een derde in Afrika. 20% van de wereldproductie is afkomstig
De Oost-Afrikaanse hooglandbananen
11
van Burundi, Rwanda, Democratische Republiek Congo, Oeganda, Kenia en Tanzanië en wordt gekweekt
Hoofdstuk 2: Banaan, een belangrijk onderdeel van de wereldeconomie
12
Productie en export van banaan
13
Groen geplukt en pas rijp in onze winkel
15
miljoen mensen in Oost-Afrika en voor 70 miljoen in West- en Centraal-Afrika, is Afrika zeer afhankelijk van
‘Gros Michel’ en ‘Cavendish’, de lievelingen van het Westen
16
de bananenteelt, zowel voor voedselzekerheid, inkomen als werkzekerheid. Toch blijven de teeltopbrengsten
Banaan: van belangrijk voedsel tot alcoholische drank, schoenen en draagzakken
17
schommelen rond 9% van hun maximale capaciteit, ondermeer wegens suboptimale landbouwcondities,
Hoofdstuk 3: Bedreigingen voor de bananenproductie loeren om de hoek
22
De Panamaziekte vernietigde nagenoeg compleet ‘Gros Michel’ en bedreigt nu ook
130 landen en een economische waarde van 44,10 miljard dollar, vormen bananen het vierde belangrijkste
voedselgewas ter wereld. De banaan vindt zijn oorsprong in Azië, maar werd lang geleden ingevoerd in Afrika, waar ze nu een belangrijke bron van voedselzekerheid geworden is. Een derde van alle bananen wordt
op velden van 0,5-4 ha groot. Slechts 15% van de wereldwijde bananenproductie wordt geëxporteerd naar
westerse landen, wat betekent dat 85% van de bananen door kleine boeren wordt geteeld voor eigen consumptie en verkoop op lokale en regionale markten. Aangezien bananen als basisvoedsel dienen voor 20
zoals droogte en gebrek aan bodemnutriënten, maar vooral door allerlei ziektes en plagen die de planten
aanvallen. Deze infecties zorgen voor een gedeeltelijke en soms zelfs totale vernietiging van de velden. De
schimmel Fusarium oxysporum is vooral een grote bedreiging voor de ‘Cavendish’-exportbananen. In Azië en
de ‘Cavendish’-banaan
23
Zuidoost-Afrika komt de productie op de grote plantages hierdoor in het gedrang en dreigt de export van
De bladvlekkenziekte veroorzaakt een sterke daling in opbrengst
24
bananen volledig stil te vallen. In Latijns-Amerika is deze schimmel nog niet aanwezig. Andere belangrijke ziek-
Virussen bedreigen de bananenteelt wereldwijd
26
De bacterie Xanthomonas is de grootste vijand van de banaan in Afrika
27
Kleine wormpjes, grote problemen
28
anderzijds in de toekomst te blijven garanderen, is er dringend nood aan verbeterde bananenvariëteiten met
De snuitkever, een onderschatte vijand?
29
verhoogde opbrengst en voedingswaarde en die resistent zijn tegen alle belagers. Momenteel worden deze
Hoofdstuk 4: Biotechnologie kan de banaan beschermen tegen zijn vijanden
30
De boer als beschermheer van zijn veld en bananenplanten
31
Wie van de drie: monocultuur, mengcultuur of gewasrotatie?
33
van de ouderplanten, het kruisen en uitpikken van de beste nakomelingen (hybriden) en het verspreiden
Zijn chemische, biologische of fysische bestrijdingsmiddelen de redders voor de banaan?
33
ervan naar de boeren minstens 17 jaar duurt. Daarnaast zullen karakteristieke eigenschappen, zoals smaak
teverwekkers zijn de schimmel Mycosphaerella fijiensis die de bladvlekkenziekte Black Sigatoka veroorzaakt, de
bacterie Xanthomonas campestris, virussen, nematoden en snuitkevers.
Om de voedselzekerheid in Afrika, Azië en Latijns-Amerika enerzijds en de wereldwijde export van bananen
verbeterde bananenvariëteiten voornamelijk via klassieke veredeling (kruisen) ontwikkeld, maar dit veredelingsproces is echter niet eenvoudig en neemt veel tijd in beslag. De meest geconsumeerde bananensoorten
zijn immers triploïd (bevatten drie sets van chromosomen), zaadloos en steriel. Men schat dat het selecteren
en geur, in de nieuwe hybriden niet altijd identiek zijn met de huidige bananenvariëteit waardoor ze niet altijd
Een werk van lange adem: het ontwikkelen van resistente bananen tegen ziekten en plagen
door de consument geapprecieerd worden. Daarom wordt geprobeerd ziekteresistenties ook via biotechno-
via kruisingsprogramma’s
35
Monnikenwerk: Veredelen van bananen in de praktijk
37
nieuwe resistente variëteiten vlugger ontwikkeld kunnen worden, maar ook dat de bananen één specifiek
Genetische transformatie van banaan: de praktijk
41
kenmerk erbij krijgen met behoud van al hun oorspronkelijke eigenschappen. Bovendien kunnen een aantal
Welke GGO-bananen bieden toekomstperspectief?
43
ziektes enkel op deze manier bestreden worden. In het geval van de bacterie Xanthomonas campestris en
Hoofdstuk 5: Banaantjes met extra vitamine A
46
6. Besluit
48
7. Referenties
50
logie en genetische bananentransformatie te introduceren. Deze technieken zorgen er niet alleen voor dat
het ‘Banana bunchy top’-virus is geen enkele variëteit gekend met een vorm van resistentie, waardoor het
inkruisen met resistentiegenen afkomstig van bananen onmogelijk is. Tegenwoordig worden de verschillende
genomen van banaan zo goed mogelijk in kaart gebracht en de verschillende eigenschappen aan hun genen
verbonden. Op die manier zal het in de toekomst mogelijk zijn om zowel met klassieke veredeling als met
biotechnologie nieuwe, verbeterde bananenvariëteiten te genereren, die bijdragen tot een duurzame, milieuvriendelijke en economisch rendabele landbouw.
Cover foto van Brigitte Uwimana, IITA
3
Cijfers en Feiten
In 2013 werd ongeveer 145 miljoen ton bananen geproduceerd in meer dan 130
landen en op meer dan 11 miljoen hectaren land.
Jaarlijks worden meer dan 1.000 miljard (bak)bananen geconsumeerd, wat banaan
wereldwijd het vierde belangrijkste voedingsgewas maakt na maïs, rijst en tarwe.
Bananen zijn een zeer belangrijk voedingsbestanddeel voor meer dan 400 miljoen
mensen. Ze staat in voor gemiddeld 15% tot 27% van hun dagelijkse calorieën.
Van de bananenproductie wordt 85% lokaal verhandeld en slechts 15%
geëxporteerd. De commerciële plantages bevinden zich hoofdzakelijk in ZuidAmerika. Export van bananen gebeurt voornamelijk naar Noord-Amerika en Europa.
De twee topproducenten van bananen, India en China, exporteren nagenoeg niets,
maar zorgen samen toch voor 35% van de wereldwijde opbrengst.
De hooglanden van Oost-Afrika produceren evenveel bananen voor lokale
consumptie als alle multinationale plantages samen voor export.
West- en Centraal-Afrika produceren 50% van alle bakbananen in de wereld.
De bananen die in het Westen geïmporteerd en geconsumeerd worden zijn het
resultaat van jarenlange selectie door de mens, die startte met de oorspronkelijke
“wilde” variëteiten die geen vruchtvlees maar enkel zaad produceerden. Nu blijven
enkel kleine, zwarte smaakloze stipjes over in de eetbare zaadloze variëteiten.
Er zijn honderden “wilde” bananenvariëteiten, maar 99% van de verkochte bananen
in de westerse supermarkten zijn genetisch identieke ‘Cavendish’-vruchten.
De schimmel Fusarium oxysporum die de commerciële productie van de ‘Gros
Michel’-variëteit volledig uitroeide, bedreigt nu ook de export van de ‘Cavendish’banaan. De meeste lokale bananenvariëteiten die in Afrika geconsumeerd worden
1
Banaan, het groene goud
van het Zuiden
zijn resistent aan Fusarium maar gevoelig voor de bacterie Xanthomonas
campestris, het ‘Banana bunchy top’-virus, nematoden en snuitkevers.
De meeste eetbare bananen zijn triploïd, steriel en zaadloos. Resistenties tegen
ziektes ontwikkelen is daarom zeer moeilijk via klassieke veredeling, maar
biotechnologie kan helpen.
Vandaag kennen we de banaan als een vlezige, zachte en zoete vrucht die bijna in iedere
fruitmand van een westers gezin voorkomt. De oorspronkelijke bananen waren echter
nauwelijks te eten en bevatten heel veel zaden. Door toedoen van menselijke selectie hebben
de huidige bananen geen of nauwelijks nog zaden, wat vooral een voordeel is bij het eten van
banaan als fruit of groente. Het ontbreken van zaden is echter ook een groot nadeel omdat
Van de Oegandese kinderen jonger dan 5 jaar kampt 73% met bloedarmoede.
bananen niet meer via zaad kunnen voortgeplant worden en dus volledig afhankelijk zijn van
Daarom worden genetisch gewijzigde bananen met verhoogde gehaltes aan vitamine
vegetatieve vermenigvuldiging. De genetische basis dan ook zeer eng geworden.
A en ijzer ontwikkeld.
Banaan, het groene goud van het Zuiden
(Figuur: Bananen in Tanzanië (Bron: R. Swennen, IITA))
5
Figuur 1.1: De struik, tros en vrucht van Musa acuminata
Figuur 1.2: De tros en vrucht van Musa balbisiana
(Bron: H. Mduma, M. Batte, R. Swennen, IITA)
(Bron: R. Swennen en M. Batte, IITA)
De voorouders van de
moderne banaan
telling. Deze soort houdt van een tropisch klimaat
zijn zeer klein en wegen slechts 2 tot 5 gram, in
ren die ontspruiten uit de wortelstam. Alle blad-
en plant zich voort zoals een doorsnee plant via
tegenstelling tot de moderne, eetbare bananen
schedes samen vormen een opeenstapeling van
zaden. Bananenplanten kunnen zich echter ook
die 200 gram kunnen bereiken. Spontane kruisin-
verticale bijna concentrische lagen. Bananenplan-
op een ongeslachtelijke manier of vegetatief
gen tussen de wilde variëteiten Musa acuminata
ten groeien op bijna alle bodems, zolang de grond
Volgens de plantensystematiek behoort de banaan
voortplanten door het vormen van zijscheuten.
en/of Musa balbisiana leidden tot zaadloze plan-
minstens 60 cm diep en niet te compact is en vol-
tot het genus Musa . Over de oorsprong van de
De Musa acuminata zorgt voor eetbare bananen
ten . Dit voor de mens zeer interessante kenmerk
doende watertransport toelaat. Daarnaast houdt
naam zijn letterkundigen het niet volledig eens: of-
die een zoete smaak hebben en een belangrijke
werd opgemerkt. Vervolgens werden specifieke
een bananenplant van warmte (27°C) en vochtig-
wel zou Carl Linnaeus de naam gegeven hebben
voedselbron zijn voor wilde dieren, zoals vleer-
kruisingsprogramma’s met de vruchtbare, “wilde”
heid (minstens 2000 mm/jaar). Er zijn echter ook
als eerbetoon aan Antonius Musa, fysicus en dokter
muizen, vogels, muizen, ratten en apen.
bananenplanten opgezet en ontstonden de bak-
variëteiten die in een gematigd klimaat kunnen
en kookbananen. Honderden zaadloze bananen-
groeien, weliswaar als sier- of vezelbanaan.
1, 2
van Keizer Augustus (eerste eeuw voor Christus), ofwel werd de naam afgeleid van het Arabische woord
De Musa balbisiana vindt zijn oorsprong in Zuid-
“Mauz”. Bijna alle moderne, eetbare bananen zijn
oost-Azië en groeit voornamelijk in ravijnen en
afkomstig van twee zadenproducerende voorou-
tropische groenblijvende bossen tot op een
ders, Musa acuminata en/of Musa balbisiana .
3
4
variëteiten werden op die manier geselecteerd .
4
Elke pseudostam vormt één bloeiwijze. Door
de zwaartekracht buigt de vruchtstengel om en
hoogte van 1100 meter. De vruchten zijn klein,
Daarom zijn de bananen krom
blauwgrijs en bevatten zeer veel zaden.
De bananenplant, die soms verkeerdelijk boom
fluweelrood is bij de exportbanaan. De typische
genoemd wordt, is een kruid, maar dan wel de
paarse kegel van de knop blijft onveranderd,
De Musa acuminata is afkomstig van de regio
aan het uiteinde vormt zich een bloemknop die
Maleisië, Indochina en Australië, maar werd ver-
Zowel M. acuminata als M. balbisiana worden niet
grootste ter wereld. De stam van de bananenplant
maar de schutbladeren vallen af en er vormen
moedelijk voor de eerste maal actief geteeld in
door de mens gegeten wegens de onverteerba-
is in feite een pseudostam of valse stam opge-
zich kleine witte bloemen in dubbele rijen. De
Papoea-Nieuw-Guinea rond 8000 voor onze jaar-
re zaden en gebrek aan vruchtvlees. De vruchten
bouwd uit de bladschedes van de bananenblade-
eerste 5 tot 15 rijen zijn vrouwelijke bloemen
Banaan, het groene goud van het Zuiden
7
Omdat de eetbare bananen geen zaden bevat-
roze. Wanneer de schil groen tot geel is, is de
bruikt in de lokale en traditionele geneeskunde7.
ten, moeten de scheuten van het onderste stam-
bakbanaan zeer zetmeelrijk en wanneer de kleur
Zo wordt de bananenschil gebruikt bij de verzor-
gedeelte door de boer geselecteerd worden om
verandert naar bruin en zwart, krijgt de bakba-
ging van brandwonden en voor de bevordering
een nieuw veld aan te planten. Verder is de bloei
naan een veel zoetere smaak en een bananen-
van wondheling. De binnenkant van de schil zou
niet afhankelijk van seizoenen waardoor het hele
geur. Bakbananen zijn doorgaans langer en heb-
de jeuk van muggenbeten verzachten en bloe-
jaar door verse bananen kunnen geoogst worden.
ben een dikkere schil dan de dessertbananen
menextracten worden gebruikt bij oogproble-
Gecultiveerde bananenplanten variëren in hoogte
die we kennen. Ze worden meestal als groenten
men. Verder worden bananen aangewend bij de
naargelang variëteit en groeicondities. De meeste
gegeten, zijn zeer voedzaam en zijn een goede
behandeling van ontstekingen, reuma, suiker-
variëteiten worden ongeveer 3-4 meter hoog, zo-
bron van kalium, vitamine A en vitamine C. Bak-
ziekte en koorts1, 7. Ze zijn ook zeer geschikt voor
als de ‘Cavendish’-banaan, maar de vorige export-
bananen zijn ook te vinden in westerse winkels.
mensen met een hoge bloeddruk, aangezien ze
zoutarm maar kaliumrijk zijn.
banaan ‘Gros Michel’ bereikt een hoogte van 7 meter. De bladeren van de bananenplant kunnen 2,7
De zoete vrucht die als fruit gegeten wordt is de
meter lang en 60 cm breed worden, maar worden
dessertbanaan. Deze is veel zoeter, heeft een
Naast de indeling volgens de uiterlijke kenmer-
door de wind gemakkelijk gescheurd.
dunnere schil en is kleiner dan de bakbananen.
ken worden bananensoorten ook volgens hun
Deze vlezige en zachte vrucht bevat geen vetten
genetische basis ingedeeld. De genetica is echter
Een gebakken banaan of eentje
uit het vuistje?
en bestaat voor 70% uit water en 27% uit sui-
complex. Bananensoorten worden gegroepeerd,
kers of koolhydraten (90 kcal/100 g). Verder is
afhankelijk van het aantal chromosomen en hun
de dessertbanaan rijk aan kalium, fosfor, calci-
oorsprong (zie kaderstuk). De meeste bananen-
um, ijzer, en de vitamines A, B6 en C. Mensen al-
soorten zijn afkomstig van de voorouders Musa
Wereldwijd wordt door kleine boeren in het Zui-
lergisch aan latex kunnen reacties krijgen na het
acuminata met het A-genoom en Musa balbisia-
den een enorme diversiteit aan bananen geteeld
eten van bananen6. Rijpe bananen bevatten ook
na met het B-genoom. De meerderheid van de
waaruit de vruchten groeien. De overige manne-
en geconsumeerd. Bananen kunnen ingedeeld
serotonine, dopamine en norepinefrine . Deze
eetbare bananen zijn triploïden en bevatten dus
lijke bloemen zijn volledig overbodig, want bana-
worden in kook-, bak-, bier- en dessertbananen,
componenten hebben een positieve invloed op
drie sets van chromosomen. Ze zijn steriel met
nen worden gevormd zonder bestuiving van de
al kan deze indeling en de interpretatie ervan
de gemoedstoestand van de mens.
AAA-genomen (dessert- en hooglandbananen),
vrouwelijke bloemen.
van plaats tot plaats verschillen. In Zuidoost-Azië
De zich ontwikkelende vruchten groeien naar het
wordt minder een onderscheid gemaakt tussen
In Zuid-Amerika, Azië, Indië en Afrika worden alle
nen) en ABB-genomen (kookbananen). Dikwijls
licht, waardoor ze zich tegen de zwaartekracht in
bak- en dessertbananen, omdat de vruchten
delen van de bananenplant, dus zowel de wor-
gebeuren er echter ook uitwisselingen tussen de
draaien, aangezien de vruchtstengel onderstebo-
daar zowel rauw als gebakken gegeten worden .
tels, pseudostam, bladeren als de bloemen ge-
A- en B-genomen.
ven hangt, en een kromme vorm aannemen. Daar-
Tenslotte bestaan er ook vezel- en sierbananen.
Figuur 1.3: Jonge tros van de ‘Cavendish’-banaan.
(Bron: FABI, Zuid-Afrika)
5
1
AAB-genomen (bak- en sommige dessertbana-
om zijn bananen dus krom. Zodra de vruchten rijp
zijn, wordt de pseudostam omgehakt en kan de
Bakbananen (of “plantains” in het Engels) komen
tros geoogst worden. Aan de basis van de plant
oorspronkelijk uit de Filipijnen en Indonesië. Ze
hebben zich echter al nieuwe scheuten gevormd
groeien nu voornamelijk in de tropische klimaten
MUSA ACUMINATA
MUSA BALBISIANA
en kunnen de dochterplanten de productie over-
van West- en Centraal-Afrika en Latijns-Amerika.
nemen, terwijl de wortelknol overblijft. De moeder-
Ze worden weleens de pasta of de aardappel van
(Aziatische banaan)
(Aziatische banaan)
plant wordt aldus vervangen, daar de scheuten of
het Zuiden genoemd, gezien ze een zeer belang-
(vruchtbaar)
(vruchtbaar)
uitlopers genetisch identiek. Door deze zijdelingse
rijk onderdeel van de dagelijkse voedselvoorzie-
scheutvorming worden bananenplantages gemid-
ning zijn. De smaak van de bakbananen verschilt
deld zes tot 30 jaar oud. De individuele planten zul-
op elk ontwikkelingsstadium en de kleur van de
len in die periode wel van plaats veranderen.
vrucht varieert van crèmekleurig, gelig tot licht-
AA
BB
AAB, ABB, AAA
etc.
(steriel)
A = 1 haploïd set van chromosomen afkomstig van Musa acuminata
B = 1 haploïd set van chromosomen afkomstig van Musa balbisiana
Figuur 1.4: Kruisingsschema voor het ontstaan van triploïde hybride bananen
Banaan, het groene goud van het Zuiden
HYBRIDE BANAAN
9
De Oost-Afrikaanse
hooglandbananen
“matoke” synoniem staat voor voedsel in Oegan-
In het gebied rond de grote meren in Afrika groeit
door vrouwen gedaan. Omdat deze vruchten
een zeer specifieke soort banaan, de Oost-Afri-
niet langdurig kunnen bewaard worden, dro-
kaanse hooglandbanaan (AAA groep). Deze
gen en vermalen de vrouwen een deel van deze
groep bestaat uit ongeveer 70 variëteiten die
bananen tot bloem, die dan als babyvoeding
onderverdeeld worden in bier- en kookbananen
gebruikt wordt.
da. Ze zijn een voorname voedselbron, vooral
in de periodes waarin andere oogsten schaars
zijn. Teelt en bereiding worden hoofdzakelijk
of matoke. Deze hooglandbananenplantages
komen voor in de regio rond het Victoriameer
In Oeganda, is “Matoke” één van de nationale
in Oeganda, in de Kilimanjaro regio’s van Tanza-
gerechten. Hiervoor worden de hooglandba-
nië, in Burundi, Rwanda en de hooglanden van
nanen geschild, gewikkeld in de bladeren en in
De kennis van het genoom, de DNA-sequentie, van banaan is zeer belangrijk. Zo kunnen genen geïdentificeerd
Oost-Congo en West-Kenia. Deze hooglandba-
een kookpot gelegd boven de stengels van een
worden die, bijvoorbeeld, fruitkwaliteit of droogteresistentie bepalen, maar ook genen die natuurlijke resistentie
nanen zijn eenvoudig te herkennen aan de tal-
bananenplant die ervoor zorgen dat het fruit
bezorgen tegen ziektes veroorzaakt door bacteriën, virussen, schimmels en nematoden. Verder wil men ook het
rijke zwarte vlekken op de pseudostam. Deze
boven het waterniveau ligt. Na een paar uren
volledige ontwikkelingsproces in kaart brengen. Bananen worden groen geoogst en pas na transport en na be-
middelgrote banaan wordt groen geoogst. Het
stomen boven een houtvuur, met af en toe toe-
handeling met ethyleen verder gerijpt. Bij dit rijpingsproces blijken nu 597 genen betrokken te zijn, die verantwoor-
vruchtvlees is wit en eerder hard, maar eerder
voeging van water, wordt het vruchtvlees zacht
delijk zijn voor het wijzigen van de celwand, of de zetmeel- en suikerconcentraties, enz.
romig en geel gekleurd eens de banaan rijp is.
en oranje. Het vruchtvlees wordt samen met de
De Oost-Afrikaanse hooglandbananen zijn zó
bladeren gepureerd en met een stukje vlees, een
belangrijk voor de plaatselijke bevolking, dat
groentensaus of gemalen pinda’s geserveerd.
DE A- EN B-GENOMEN IN DETAIL BEKEKEN
In de Musa familie zijn er vier genomen aanwezig die alle afkomstig zijn van wilde variëteiten: Musa acuminata
[A-genoom, diploïd (2n = 2x) met 22 chromosomen] , Musa balbisiana [B-genoom, 2n = 2x = 22 chromosomen],
8
Musa schizocarpa [S-genoom, 2n = 2x = 22 chromosomen], en Musa textilis [T-genoom, 2n = 2x = 20 chromosomen]8. Aangezien de meeste bananen echter kruisingen zijn van de wilde diploïden Musa acuminata en Musa
balbisiana9-11, zijn verschillende combinaties van de A- en B-genomen12 ontstaan.
In 2012 werd de sequentie van het A-genoom van de DH-Pahang cultivar bepaald13. De DH-Pahang banaan is een
homozygoot of dubbele haploïd (AA) van het M. acuminata genotype. Eén haploid genoom telt 11 chromosomen,
dus 2n telt 22 chromosomen. Van dit A-genoom werd 90% van de sequentie bestaande uit 523 megabasen bestudeerd. Men identificeerde 36.542 genen die coderen voor een eiwit. Daarnaast bestaat de helft van het A-genoom
uit transposons of springende genen13.
Eén jaar later, in 2013, werd ook het B-genoom ontrafeld8. Voor deze sequentieanalyse werd het diploïde genoom
van de Musa balbisiana cultivar Pisang Klutuk Wulung gebruikt. Deze cultivar vertoont gedeeltelijke resistentie
tegen de “black leaf streak” schimmel. Via de sequentie wil men de resistentiegenen identificeren. Dit B-genoom is
21% kleiner dan het A-genoom en bevat 36.638 genen die coderen voor eiwitten, wat ongeveer overeenkomt met
hetzelfde aantal genen als het A-genoom. Het B-genoom draagt voornamelijk bij tot de sterkte van de plant en
tolerantie voor biotische en abiotische stressfactoren.
Banaan, het groene goud van het Zuiden
Figuur 1.5: De Oost-Afrikaanse hooglandbanaan
Figuur 1.6: Matoke bananen na stomen
uit Oeganda. (Bron. R. Swennen)
(Bron: R. Swennen)
11
Productie en export van banaan
bron van inkomen. In Oeganda, bijvoorbeeld, zijn
In 2013 werden meer dan 145 miljoen ton bana-
naan voor voedsel, inkomen en werk.
meer dan 13 miljoen mensen afhankelijk van ba-
nen geteelt in meer dan 130 landen op een oppervlakte van meer dan 11 miljoen hectare14 (Ta-
De AAA-groep ‘Cavendish’-banaan is de meest
bel 1, Figuur 2.2). De topproducenten zijn India,
geteelde soort en is goed voor 28% van de lo-
met ongeveer 27 miljoen ton (overeenkomend
kaal geconsumeerde vruchten. Daarnaast wordt
met 19% van de totale productie), en China met
vooral de AAB-subgroep van bakbananen ge-
12 miljoen ton (8% van de totale productie). Oe-
teeld voor consumptie in West-Centraal-Afrika
ganda is de derde grootste producent met on-
en Zuid- en Centraal-Amerika (21%). Met andere
geveer 9,5 miljoen ton (kook- en bier)bananen,
woorden, twee van de 50 erkende subgroepen
gevolgd door de Filipijnen (8,6 miljoen ton) en
zijn verantwoordelijk voor meer dan 60% van de
Brazilië (6,9 miljoen ton).
totale productie15.
Opmerkelijk is dat de grootste producenten In-
Ondanks de grote diversiteit, wordt de exportpro-
dië en China nagenoeg geen bananen uitvoe-
ductie volledig gedomineerd door de ‘Cavendish’
ren. Van de wereldwijde productie werd in 2012
bananen (zie verder), een groep zoete bananen-
slechts ongeveer 15% (of 20 miljoen ton) geëx-
variëteiten die in het veld heel sterk op elkaar ge-
porteerd . De overgrote meerderheid van de
lijken en eens verpakt niet te onderscheiden zijn.
bananen wordt geteeld voor eigen consumptie
Commerciële bananenplanten worden vaak op
of lokale verkoop op het land of in de tuintjes
grote plantages geteeld. Bijna 70% van de vol-
van kleine boeren. Door de zeer grote verschei-
ledige exportmarkt is in handen van slechts 10
denheid aan variëteiten blijft de rijke diversiteit
landen. De grootste uitvoerder is het Zuid-Ame-
behouden. Bananen kunnen het hele jaar ge-
rikaanse land Ecuador14 (Figuur 2.2, Tabel 1) met
oogst worden, waardoor ze een constante bron
ongeveer 5,2 miljoen ton bananen of 26% van de
van voedsel en voor de kleine boer een continue
totale export. Ecuador produceert ongeveer 6,5
14
miljoen ton bananen, wat betekent dat ongeveer
80% van de productie geëxporteerd wordt. De
2
Filipijnen met ongeveer 2,6 miljoen ton of 14%
Banaan, een belangrijk onderdeel
van de wereldeconomie
van de totale exporthoeveelheid zijn de tweede
grootste uitvoerder. De top vijf wordt vervolledigd met Guatemala (2 miljoen ton of ongeveer
10%), Costa Rica (1,9 miljoen ton of ongeveer
10%) en Colombië (1,8 miljoen ton of ongeveer
9 %). Voor commerciële doeleinden worden dessertbananen hoofdzakelijk in Zuid-Amerika en
de Caraïben gegroeid, kookbananen in Azië en
bakbananen in Centraal- en West-Afrika en in
Zuid- en Centraal-Amerika4. In 2013 waren vijf
bedrijven verantwoordelijk voor 44% van de in-
(Figuur: Geoogste ‘Cavendish’-bananen in Suriname (Bron: R. Swennen))
Banaan, het groene goud van het Zuiden
Figuur 2.1: Exportbananen (‘Cavendish’) worden ingepakt voor transport
in Zuid-Afrika (Bron: FABI, Zuid-Afrika)
13
%
do
r2
6,
39
%
,03
%
ua
Ec
9,
ss
jk 5,7
1%
65
ë
bi
om
l
Co
isc
he
Fe
7,11%
de
ra
tie
6,7
6%
Veren
igd K
onikri
8,
Ru
%
en
3%
2%
2,8
%
ada
3,1
Can
k
rij
nk
a
Fr
3,
17
ijn
3,4
2%
lip
%
31
n1
%
de Belgen gemiddeld 8 kg.
2,5
België
ijne
,44
nenimport. Na het rijpingsproces worden 94%
na
Ga
Fi
6%
d6
inwoner, de Britten 12,5 kg, de Fransen 8,5 kg en
9%
Filip
a 6,5
an
Antwerpen is de belangrijkste haven voor bana-
%
24
itsl
den eten ongeveer 19 kg bananen per jaar per
,72
%
Japan 5,83
elk een import van meer dan één miljoen ton.
and
8%
10,0
woners van Noord-Europa geliefkoosd. De Zwe-
Oeg
%
land en Japan vervolledigden de top vijf met ook
VS
4%
4,76
alle gegeten vruchten en zijn vooral door de in-
2%
,9
30
China 8,5
zilië
de Russische federatie 1,26 miljoen ton. Duits-
re
Du
vertegenwoordigen bananen ongeveer 11% van
%
ala
tem
Gua
miljoen ton bananen, België 1,32 miljoen ton en
de
1%
Ecuador 4,52%
in China en 3 kg in Rusland. In de Europese Unie
,22
34,3
Bra
porteerde de Verenigde Staten ongeveer 4,6
An
n
12 kg in de Verenigde Staten, 8 kg in Japan, 5 kg
31
3,4
voor in de lijst van importlanden. In 2012 im-
re
lië
De jaarlijkse consumptie per inwoner bedraagt
ere
de
Ira
Vanzelfsprekend komen totaal andere landen
And
Import
Ita
de en geconsumeerde vrucht ter wereld te zijn.
An
ica 9
,57%
Italië). De banaan blijkt de meest geëxporteer-
Export
Cos
ta R
verkochten (www. bananalink.org.uk).
Productie
19
in Duitsland, 12% in Groot-Brittannië en 8% in
Ind
ië
landen van de Europese Unie terecht komt (18%
grote bedrijven of direct aan de kleinhandelaar
ië 3,73
%
kleinere bedrijven die hun vruchten ofwel aan de
Colom
b
van 1,3 miljard Euro, waarvan 26% in andere
3,
61
%
esië
3,7
%
overige 56% werd verhandeld door nationale
Figuur 2.2: Overzicht van de wereldwijde productie, export en import van (bak)bananen.
on
pen terug geëxporteerd, met een totale waarde
m
er
oe
n
(11%), Dole (11%), Fyffes (6%) en Noboa (2%). De
Ind
van de geïmporteerde bananen vanuit Antwer-
Ka
ternationale handel: Chiquita (13%), Del Monte
ment moet echter ook rekening gehouden worden met het transport van veld naar consument.
Rijp geoogst fruit is zachter dan onrijp fruit en
Tabel 1: Overzicht van de wereldwijde productie, export
wordt dus gemakkelijker beschadigd tijdens het
en import van (bak)bananen
transport. De houdbaarheid van rijp geoogste
bananen is ook belangrijk en bedraagt amper 7
Land
Productie
(Miljoen ton)
in 2013
Land
Indië
27,6
China
Export
(Miljoen ton)
in 2012
Land
Ecuador
5,2
Verenigde Staten
4,6
12,4
Filipijnen
2,6
België
1,3
Oeganda
9,5
Guatemala
2
Russische Federatie
1,3
Filipijnen
8,6
Costa Rica
1,9
Duitsland
1,2
Brazilië
6,9
Colombië
1,8
Japan
1,1
Ecuador
6,5
Verenigd Koninkrijk
0,6
Colombië
5,4
Italië
0,6
Indonesië
5,4
Iran
0,6
Kameroen
5,2
Frankrijk
0,6
Ghana
3,8
Canada
0,5
Totale productie
145
Totale import in de
19,7
Import
(Miljoen ton)
in 2012
tot 10 dagen, wat te kort is om de lange afstand
tussen de tropische gebieden tot aan de westerse consument te overbruggen. Daarom worden
in de wereld
Totale export in
de wereld
19,7
wereld
Aangezien niet alle landen een duidelijk onderscheid maken tussen de hoeveelheid geproduceerde bananen en
bakbananen, werden hier totale hoeveelheden opgeteld14.
Banaan, het groene goud van het Zuiden
exportbananen steeds onrijp en groen geplukt,
worden de vruchten voorzichtig behandeld en
koel bewaard (13,5 tot 15°C) en wordt de duur
Figuur 2.3: De exportbananen worden voor
van hun vervoer zo kort mogelijk gehouden. Men
verpakking groen geoogst. (Bron: R. Swennen)
wil namelijk voorkomen dat het hormoon ethyleen, dat aangemaakt wordt door de rijpende
Groen geplukt en pas rijp in
onze winkel
vrucht, het rijpingsproces in gang zet.
Exportbananen zijn eerst groengeel van kleur
bij een temperatuur van 17°C. De geïmporteer-
om daarna naar bruingeel te verkleuren tijdens
de bananen worden zo dicht mogelijk bij de con-
het rijpen. Net zoals iedere vrucht is de smaak en
sument, in het importerende land, gerijpt. De
de textuur van een aan de plant gerijpte banaan
mooie gele kleur van de bananen in de wester-
het best. Bij het bepalen van het ideale oogstmo-
se supermarkten wordt dus bekomen door een
Na het transport worden de bananen in speciale
kamers gerijpt die met ethyleen gevuld worden
15
clamecampagnes werden zelfs opgezet om aan
mel, de ‘Cavendish’. De ‘Cavendish’-banaan werd
onale markten. Bananen en bakbananen blijken
te geven dat bananen voor iedereen, ook baby’s,
in Vietnam gevonden. Het is ook een triploïde ba-
immers meer mensen per oppervlakte-eenheid
goed waren en dat ze op elk moment van de dag
naan van de AAA groep en werd voor het eerst
te voeden dan om het even welk ander gewas19.
konden gegeten worden.
geteeld in 1836. De ‘Cavendish’ plant wordt 3 tot
Bovendien is de productie veel goedkoper dan
5 meter hoog en de vruchten zijn minder zoet en
die van rijst en tarwe20 en is de teelt minder ge-
Tot in de jaren 1950 was ‘Gros Michel’ een zeer
kleiner dan die van de ‘Gros Michel’ planten. Sinds
voelig voor wereldwijde prijsschommelingen,
belangrijk exportproduct, vooral naar de Ver-
de jaren 1960, na het verdwijnen van de ‘Gros Mi-
wat wel het geval kan zijn voor rijst, maïs en tar-
enigde Staten en Europa, maar werd reeds sinds
chel’, nam de ‘Cavendish’ de rol van exportbanaan
we19. Inderdaad, de banaan is wereldwijd een
Figuur 2.4: ‘Gros Michel’ bananen op de markt
het begin van de jaren 1900, bedreigd door de
over. Sindsdien behoren alle bananen die voor de
heel belangrijk basiselement in de voeding, met
in Tanzanië (Bron: R. Swennen)
schimmel Fusarium oxysporum (zie Hoofdstuk
wereldwijde handel gebruikt worden tot de ‘Ca-
een gemiddelde van 21 kg/inwoner/jaar. Deze
3). Deze schimmel dook voor het eerst op in
vendish’ variëteit, die eveneens geen zaden bezit
gemiddelden liggen echter veel hoger in ontwik-
geïnduceerd rijpingsproces, dat de smaak en de
Panama en verspreidde zich vlug over heel de
en dus onvruchtbaar is en via vegetatieve voort-
kelingslanden. In Oeganda, Burundi en Rwanda
textuur van de banaan bepaalt. De behandeling
wereld17, 18. De totale productie van ‘Gros Michel’
planting vermeerderd moet worden. Hierdoor zijn
eet men tot 300-400 kg bananen/inwoner/jaar,
blijkt essentieel te zijn want groene bananen die
viel stil in Zuid- en Centraal-Amerika en Afrika en
alle ‘Cavendish’-bananen genetisch identiek en zijn
wat neerkomt op een gemiddelde van 3-11 bana-
nooit met ethyleen behandeld worden zullen
nagenoeg alle grote telers gingen failliet. Tegen-
de vruchten die in de jaren 1960 verkocht werden
nen/dag/persoon21. Bijna elke maaltijd bestaat uit
niet volledig rijpen voordat ze rotten.
woordig wordt de ‘Gros Michel’ nauwelijks nog
in het Westen identiek aan deze die we vandaag in
gepureerde bananen die gemengd worden met
geteeld, enkel in delen van Thaïland en door
de supermarkt vinden.
kruiden. Studies toonden aan dat in landen zoals
‘Gros Michel’ en ‘Cavendish’,
de lievelingen van het Westen
kleine boeren in Afrika, de Caraïben en Zuid- en
Voor de exportindustrie worden sinds de jaren
exportbanaan, kwam een nieuwe variëteit op de
1870 slechts twee variëteiten geteeld, ‘Gros Mi-
markt die wel bestand bleek te zijn tegen de schim-
Centraal-Amerika.
In de periode dat de ‘Gros Michel’ verdween als
Oeganda, Rwanda en Burundi, bananen verant-
Banaan: van belangrijk voedsel
tot alcoholische drank,
schoenen en draagzakken
woordelijk zijn voor 30% van de calorieopname,
en dat dit in bepaalde gebieden zelfs kan oplopen
tot 60%22. In Oeganda zou er geen hongersnood
heersen dankzij de teelt van bananen.
chel’ en ‘Cavendish’ . Gros Michel, ook wel ‘Big
Uit de productie- en exportcijfers is heel duide-
Bananen kunnen op verschillende manieren ge-
Mike’ genoemd, is een triploïde variëteit van de
lijk dat het overgrote deel van de in het Zuiden
geten worden: gebakken, gestoomd, gekookt of
AAA groep, met als voorouder Musa acuminata.
geproduceerde bananen bedoeld zijn voor eigen
rauw. Daarnaast worden ze ook verwerkt in jam,
consumptie en verhandeling op de lokale en regi-
bier, koekjes en beschuiten. In Leuven (België)
16
‘Gros Michel’ werd al in de jaren 1800 gecultiveerd en verhandeld. Het is een zeer zoete en
smaakvolle banaan met een meer romige structuur dan de huidige commerciële banaan. De
plant kan tot 7 meter hoog groeien en de vruchten zijn ongeveer 18 tot 23 cm lang. Het is de
lieveling van de producenten omdat deze vrucht
een zeer sterke, dikke schil bezit waardoor hij
weinig extra zorg nodig heeft tijdens groei en
transport. Verder vertonen deze planten zeer
hoge opbrengsten. Enkele rijke Amerikanen
kwamen op het idee om een handel in bananen
Figuur 2.5: Lokale handel in “Sukali nduzi”
op te zetten. De consumptie steeg enorm en re-
bananen in Tanzanië (Bron: R. Swennen)
Figuur 2.6: (A) Bananenlikeur (www.musaLova.be); (B) Bananenbier (Tanzanië) (Bron: R. Swennen); (C) Tonto Bananenbier
(Oeganda) (Bron: Y. Lokko, Unido); (D) Bananenchips (Bron: D. Amah, IITA) en (E) Gefrituurde bananen (Bron: R. Swennen)
Banaan, het groene goud van het Zuiden
17
Figuur 2.7: (A) Dakbedekking gemaakt van bananenbladeren (Oeganda) (Bron: R. Swennen) , (B) Kledij gemaakt van bananenvezels (Tanzanië) (Bron: M. S.R. Byabachwezi, ARI Maruku, Tanzanië)
bananenplant uit de ondergrond aan de opper-
planten verwacht worden. Daarnaast hebben
vlakte worden gebracht. De afgevallen cacaobla-
de bananenplanten ook een belangrijke ecologi-
deren houden de grond onkruidvrij en vochtig.
sche functie. Ze verminderen de landerosie en
Dankzij deze symbiose tussen cacao- en bana-
de oogstoverschotten geven na elke productie-
nenplant kan een grotere opbrengst van beide
cyclus ook voedingsstoffen terug aan de grond24.
DE MEERWAARDE VAN NEVENPRODUCTEN VOOR DE KLEINE
BOEREN BIJ DE BANANENTEELT IN OEGANDA
Bananen (bak-, kook- en dessertbananen) zijn een zeer belangrijk voedingsgewas in Oeganda en bieden voedsel-
ontwikkelde men “Musa Lova”. Ook bananenpan-
zels gemaakt . In sommige gebieden, en vooral
zekerheid aan ongeveer 13 miljoen mensen. Over het algemeen worden de bananen door kleine lokale boeren ge-
nenkoeken zijn zeer populair in Zuid- en Zuidoost
in het droogteseizoen, vormen bananenblade-
produceerd en volledige trossen onmiddellijk op de plaatstelijke markt aangeboden. Hierdoor geraken de markten
Azië. Onrijpe bakbananen worden hoofdzakelijk
ren en versneden stengels een belangrijk be-
dikwijls oververzadigd en worden de bananen ook naar stedelijke gebieden getransporteerd. Een klein deel van de
gebruikt om bananenchips te maken. Dit zijn dun-
standdeel van het veevoeder.
bananen vindt zelfs zijn weg naar Europa, de Verenigde Staten en naburige landen zoals Zuid-Soedan, waar de
23
voedselprijzen meestal hoger liggen.
ne uitgedroogde bananenschijfjes met een donkere kleur en een uitgesproken bananensmaak.
De bananenplant heeft in bepaalde streken van
Bananensap maken is door de textuur van de
Afrika, zoals Ivoorkust, nog een andere belang-
Tijdens opslag en transport zijn de bananen echter zeer gevoelig voor overrijping en rotting, waardoor de boeren,
vrucht heel moeilijk aangezien het extractie ver-
rijke functie. De bananenplanten worden als
die al zeer weinig verdienen, belangrijke inkomsten verliezen en er een tekort aan voedsel dreigt. Daarom is het
eist, maar de vruchten worden wel aangetroffen
schaduwplanten voor de cacao- of koffieplanten
verwerkingsproces van bananen zeer belangrijk, niet alleen om de houdbaarheid van de vruchten te verlengen,
in bloem, soep, ontbijtgranen en zelfs in indus-
geteeld . Beide planten halen hun voordeel uit
maar ook om bijproducten van het gewas te produceren, waardoor nieuwe markten ontstaan. Hierdoor krij-
triële versies van fruitsalades en smoothies. In
deze gezamenlijke teelt: wanneer de cacaoplant
gen bananen een toegevoegde waarde, verhoogt de voedselzekerheid en krijgen de boeren hogere inkomsten.
Zuid- en Zuidoost-Azië wordt ook het hart van de
nog jong is, zorgt de schaduw van de bananen-
Bananen worden momenteel verwerkt tot bv. bier (Tonto) en lokale gin (Waragi). Verder worden in supermarkten
bananenbloemen, dat een beetje aan de artisjok
plant voor bescherming. De cacaoplant, waar-
nieuwe producten aangeboden zoals vacuüm verpakte geschilde matoke bananen, bananensap, -wijn en -meel,
doet denken, dikwijls gegeten, want ook hier zijn
van de wortels veel dieper in de bodem groei-
chips en gedroogde vruchten. Het sap uit bananen halen kan zowel op een moeilijke mechanische als op een
de vlezige delen van de schutbladeren en het hart
en, zorgt ervoor dat voedzame stoffen voor de
enzymatische manier gebeuren. Hoewel de enzymatische werkwijze veel efficiënter is en er hogere hoeveelheden
24
van de bloem eetbaar. In Oost-Afrikaanse landen
sap verkregen worden, draagt de traditionele extractiemethode toch de voorkeur weg. Men schat dat er jaarlijks
zoals Burundi, Rwanda en Oeganda worden kook-
2 miljoen liters waragi/alcohol en 1,4 miljoen bier verkocht worden.
bananen hoofdzakelijk gebruikt in puree, sap,
Hoewel de meeste bijproducten van banaan op kleine schaal geproduceerd worden, bestaan er toch een aan-
bier, gin, wijn en likeur.
tal bedrijven die zich gespecialiseerd hebben in het produceren van hoogwaardige producten. Het Presidentiële
In de tropische landen worden bananenblade-
Initiatief voor Industriële Bananenontwikkeling (PIBID) produceert het bananenmeel “Tooke floor”. “Afri Banana
ren gebruikt als verpakkingsmateriaal, dakbe-
Products Ltd” is een bedrijf dat vacuumverpakte bananen, bananensap en –wijn produceert, “Forest Fruit Foods
dekking, paraplu’s en zelfs als wegwerpbordjes.
Ltd” is een KMO (kleine of middelgrote onderneming) dat bananensap maakt en “Fruits of the Nile” ondersteunt
De vezels van de bananenbladeren worden ver-
kleine boeren bij de productie van gedroogde vruchten voor export.
werkt in handtassen, schoenen, papier, touw,
manden, theebuiltjes en bankbiljetten. De kimo-
Figuur 2.8: Het samen opgroeien van bananenplanten
Ook de afvalproducten op de bananenplantages hebben een belangrijke meerwaarde. Zowel bananenbladeren,
no’s in Japan werden jarenlang van bananenve-
en koffieplanten in Tanzanië (Bron: R. Swennen)
vezels, schillen als pulp worden aangewend voor de aanmaak van nieuwe producten. De vezels worden gebruikt
als dakbekleding en voor het maken van kabels voor het transport van thee. Bananenbladeren hebben een sterke
Banaan, het groene goud van het Zuiden
19
culturele betekenis in Oeganda en dragen bij tot de specifieke smaak van bepaalde maaltijden. Bananenschillen
en -pulp en bananen die niet geschikt zijn voor verkoop en/of menselijke consumptie worden verwerkt in dierenvoeder. Recent ontwikkelde het Oegandees Industrieel Onderzoeksinstituut (IRRI) een proces voor de extractie
van vezels uit de pseudostam voor de aanmaak van papier en textiel. Tenslotte kan het afval van bananen ook
aangewend worden voor de productie van biogas, waardoor men voor de energiebehoeften minder afhankelijk
wordt van brandhout en houtskool. Het slib dat tijdens de productie van biogas als bijproduct gevormd wordt
kan als compost gebruikt worden.
Door middel van financiële steun van de “Global Environmental Facility” (GEF) en het “Least Developed Country
Fund (LDCF), staat UNIDO de Oegandese regering bij tijdens de ontwikkeling van een programma om duurzame producten afkomstig van de bananenteelt te ontwikkelen. Het project wil vooral de kwetsbare populaties in
West-Oeganda beter leren omgaan met de gevolgen van de klimaatsveranderingen en de invloed op de productie
van bananen en hun bijproducten om zo meer inkomsten te kunnen genereren, armoede te bestrijden en voedselveiligheid te garanderen.
Figuur 2.9: Bananencollectie in Leuven (Bron: R. Swennen)
Figuur 2.10: Bananenweefsel wordt ook
bewaard in vloeibare stikstof (Bron: B.
Panis, Bioversity International)
soorten. Eén keer per jaar worden alle zaadkiemen in proefbuisjes overgeënt en in een koele (15°C) kamer met
zwakke lichtsterkte geplaatst om trage groei te verzekeren. Vanuit Leuven zijn de voorbij 30 jaren bananenstekjes
vertrokken naar meer dan 355 plaatsen in 100 verschillende landen. De plantjes worden dan uit hun winterslaap
DE GROOTSTE BANANENCOLLECTIE BEVINDT ZICH IN LEUVEN
gehaald om nieuwe bananenplanten via weefselcultuur aan te maken, die vervolgens in kweekkamers bij 27°C
België voert al meer dan 100 jaar onderzoek naar bananen. Edmond De Langhe, Professor emeritus aan de
gen zijn 75% bestemd voor lokale onderzoekers en 25% voor telers, NGO’s en landbouworganisaties. De plantjes
Katholieke Universiteit (KU) Leuven startte in de jaren 1950 met het bestuderen en verzamelen van bananen.
worden gratis aangeboden, maar de afnemer mag ze niet patenteren. Na de genocide in Rwanda en bijhorende
Sinds 1976 ondersteunde de Belgische overheid het onderzoek naar bakbananen bij de “International Institute of
massale volksverhuizing vanuit Rwanda en Burundi naar Tanzanië, werden 70.000 plantjes naar Tanzanië opge-
Tropical Agriculture” (IITA). In 1979 vervoegde doctoraalstudent Rony Swennen het IITA team en zorgde hij mee
stuurd die ter plaatse vermenigvuldigd werden en 6 miljoen gezonde bananenplanten leverden. Hierdoor zagen
voor de uitbouw van een onderzoeksprogramma. Momenteel beheert Prof. Dr. Rony Swennen, directeur van het
de kleine boeren hun opbrengsten en inkomen verdrievoudigen en werd hongersnood vermeden.
geplaatst worden tot ze klaar zijn voor verzending. Elke aanvrager krijgt vijf stalen per soort. Van deze verzendin-
laboratorium voor Tropische Plantenteelt aan de KU Leuven, de grootste bananencollectie ter wereld. Ook ligt het
laboratorium van Edmond De Langhe en Rony Swennen aan de oorsprong van het “International Network for the
De onderzoekers in Leuven houden verder van elke soort de gegevens over teeltomstandigheden, eigenschappen
Improvement of Banana and Plantain” (INIBAP), een wereldwijde samenwerking die waakt over de biodiversiteit
en verschillende milieus waarin ze zich ontwikkelen bij. Verder wil men alle bananensoorten ook bij -196°C in
van de banaan en in 2015 zijn dertigste verjaardag vierde. Het INIBAP, nu deel van “Bioversity International”, is
vloeibare stikstof bewaren om op deze manier ook de biodiversiteit te kunnen verzekeren. Momenteel zijn reeds
een netwerk dat in Afrika, Azië, en Latijns-Amerika werkzaam is. In elk van deze continenten krijgt het netwerk
700 soorten diepgevroren, maar het duurt zeker nog 12 jaar om ook de overige 800 in te vriezen. Deze techno-
steun van regionale en nationale netwerken, bestaande uit leden van boerenorganisaties, NGO’s, coöperaties,
logie (cryopreservatie) past deze onderzoeksgroep niet alleen toe op banaan, maar ook op 27 plantenfamilies
universiteiten en privéondernemingen.
waaronder dadelpalmen, zoete aardappelen, witloof, suikerbieten, cassave en aardbeien. Daarnaast werd de
bananencollectie ook gedupliceerd om zeker te zijn dat ze nooit verloren kan gaan, bijvoorbeeld bij brand. Het
De Leuvense bananencollectie staat onder toezicht van de Verenigde Naties en telt momenteel meer dan 1500
Laboratorium voor Tropische Plantenteelt, IITA en Bioversity International werken ook actief samen met de kleine
ziektevrije bananensoorten, waarvan het aantal nog steeds aangroeit. Er bestaan in het wild immers nog een
boeren in Afrika, vooral om ze te overtuigen om meerdere bananenvariëteiten tegelijk te kweken en aldus de kans
groot aantal onbekende bananensoorten en regelmatig trekken onderzoekers de jungle in op zoek naar nieuwe
op een geslaagde oogst te verhogen.
Banaan, het groene goud van het Zuiden
21
De Panamaziekte vernietigde
nagenoeg compleet
‘Gros Michel’ en bedreigt nu
ook de ‘Cavendish’-banaan
dere planten. Doordat deze schimmel de plant
eerst volledig inwendig aantast, wordt de ziekte
zeer laat opgemerkt. De besmetting en verspreiding naar andere planten zijn dan dikwijls te ver
gevorderd om nog te kunnen ingrijpen.
De sporen van Fusarium oxysporum zijn langer
De Panamaziekte wordt veroorzaakt door de
dan een jaar kiemkrachtig en kunnen 20 tot zelfs
grondschimmel Fusarium oxysporum f. sp. cubense
40 jaar in de aarde overleven, zelfs in afwezig-
. Deze grondschim-
heid van de gastheer (banaan)28, 29. De Panama-
mel infecteert de bananenplant via de wortels en
ziekte Foc-TR1 heeft vooral de ‘Gros Michel’ vari-
verspreidt zich via de sapstroom in het geleidings-
ëteit aangetast waardoor zijn commerciële teelt
weefsel doorheen de volledige bananenplant. Op
sinds 1960 volledig verdwenen is. Al in de jaren
verschillende plaatsen in de vaatbundels vormt de
1960 beschouwde men de Panamaziekte als één
schimmel een soort gel die de doorvloeiing van
van de meest vernietigende plantenziektes. De
vloeistoffen en nutriënten doorheen de plant be-
economische schade werd toen geraamd op 2,3
lemmert, met uitdroging van de plant als gevolg .
miljard dollar en vele telers gingen failliet. De
Wanneer de pseudostam van een geïnfecteerde
schimmel blijkt echter ook dodelijk te zijn voor
plant wordt opengesneden, blijken de vaatbundels
een hele reeks lokale bananenvariëteiten en
volledig bruin/zwart te zijn. Andere symptomen
vormt dus ook een bedreiging voor de bananen-
zijn vergeelde bladeren en wortel- en knolrot. De
teelt in ontwikkelingslanden.
(Foc), Tropical Race 1 (TR1)
18, 25, 26
3
Bedreigingen voor de
bananenproductie loeren om de hoek
De bananen zoals wij die vandaag in de winkels vinden worden vooral geteeld op grote
plantages. Door het ontbreken van zaden resulteren de bananenplanten nagenoeg allemaal
van vegetatieve voorplanting, waardoor ze genetisch identiek zijn en dezelfde positieve,
maar ook negatieve, eigenschappen bezitten. De volledige teelt kan hierdoor zeer gevoelig
zijn voor aanvallen van bacteriën, virussen, schimmels, nematoden en insecten wanneer
deze bananenplanten niet de juiste resistenties bezitten. Ook de bananensoorten die door
de kleine boeren voor lokale consumptie geteeld worden, zijn vaak genetisch identiek en
zeer gevoelig voor mogelijke belagers. Eenmaal een bepaalde bananensoort bedreigd is
door een ziekte, gaat de verspreiding van de ziekte en vernietiging van de oogst dan ook
zeer snel.
(Infectie van bananenplanten met Fusarium oxysporum (Bron: A. Viljoen, Stellenbosch Universiteit, Zuid-Afrika))
Banaan, het groene goud van het Zuiden
27
ziekte werd voor het eerst waargenomen begin
1900 in Panama, vandaar de naam, en verspreidde
Aangezien de ‘Gros Michel’ variëteit niet meer
zich van daaruit naar naburige landen.
grootschalig kon geteeld worden, ging men op
zoek naar een andere commerciële banaan, de
Schimmels planten zich onder meer voort door
‘Cavendish’ variëteit (zie Hoofdstuk 2). De ‘Ca-
middel van sporen (microscopische overlevings-
vendish’ plant bleek immuun te zijn tegen de
en voortplantingsstructuren). Deze schimmel-
schimmel Fusarium oxysporum Race 1 en werd
sporen komen via de afgestorven plant terug in
vanaf de jaren 1960 steeds belangrijker voor de
de bodem terecht en infecteren vervolgens an-
handel. Aldus geraakte de infectieziekte wat op
23
delijk ook al aanwezig in andere gebieden van
Zuidoost-Azië36. Omdat geïnfecteerde bananenbladeren heel dikwijls als verpakkingsmateriaal gebruikt werden, verspreidde deze ziekte
zich heel vlug buiten Azië. De schimmelsporen
worden ook zeer vlug via de wind of regen verspreid36. In 1972 werd de ziekte een eerste
maal waargenomen in Honduras en vervolgens
in Belize, Mexico, Guatemala, El Salvador, Nicaragua, Costa Rica, en vele andere Centraal- en
Figuur 3.1: Bananenplantage aangetast door de panamaziekte
Figuur 3.2: Dwarse doorsnede van de pseudostam na infectie
(Bron: A. Viljoen, Stellenbosch University, Zuid-Afrika)
met de Fusarium schimmel (Bron: A. Viljoen)
Zuid-Amerikaanse landen37. Bijna gelijktijdig, in
1973, dook deze ziekte ook voor het eerst op in
het Afrikaanse land Zambia, maar komt het nu al
voor in gans Sub-Sahara-Afrika.
de achtergrond, al werd in 1986 gewaarschuwd
Fusarium oxysporum grondschimmels versprei-
dat nieuwe infectieziektes dreigden uit te breken
den zich voornamelijk via vervuilde grond en
De schimmel Mycosphaerella fijiensis ontwikkelt
door het telen van zoveel genetisch identieke
water, maar ook via schoeisel, kledij, gereed-
sporen op het bananenblad bij hoge vochtig-
schap, insecten, irrigatiewater en geïnfecteerde
heid en een temperatuur van ongeveer 27°C36.
stekken35. Wanneer plantgoed afkomstig is van
Een twee- tot drietal dagen na infectie dringen
In 1992 werd inderdaad een nieuwe zeer agres-
weefselcultuur, kan men met ziektevrije planten
de sporen het bananenblad binnen via de huid-
sieve variant van de Fusarium stam ontdekt: Fusa-
werken (zie Hoofdstuk 4), maar ze kunnen ech-
mondjes*en vermenigvuldigen ze zich verder.
rium oxysporum Tropical Race 4 (Foc-TR4) die voor
ter heel vlug aangetast worden na beplanting in
De infectie start aan de onderzijde van het blad
het eerst waargenomen werd in Azië op het ein-
geïnfecteerde gronden. Het is nu bang afwach-
en vormt kleine zwarte vlekjes die zich geleidelijk
de van de vorige eeuw en nu ook al voorkomt in
ten wanneer deze Foc-TR4 stam Centraal- en
verder uitbreiden en parallel met de bladnerven
Taiwan, Indonesië, Maleisië, de Filipijnen, China en
Zuid-Amerika zal bereiken en hoe vlug de ver-
zwarte strepen vormen, waar geen fotosynthe-
Noord-Australië. In deze gebieden is de teelt van
spreiding van deze ziekte in Amerika en Afrika
se** meer kan doorgaan38. De energieproduc-
tie met meer dan 40% door deze ziekte24. In de
de ‘Cavendish’-banaan sterk gedaald, wat proble-
de ‘Cavendish’ productie zal bedreigen, met een
tie vermindert en het blad sterft uiteindelijk af.
jaren 1970 produceerde Oeganda nog 15 tot 20
matisch is als geweten is dat China, de Filipijnen
wereldwijde impact op de internationale handel.
De gereduceerde energietoevoer veroorzaakt
ton bananen per hectare, maar in gebieden waar
een vervroegde rijping van het fruit waardoor
deze schimmel aanwezig is, bedraagt de produc-
De bladvlekkenziekte veroorzaakt
een sterke daling in opbrengst
het ongeschikt wordt voor export. Normaal be-
tie momenteel nog slechts 6 ton per hectare.
waarvan al 80% van de plantages besmet zijn .
De bladvlekkenziekte of “black leaf streak“ of
ontwikkelen39. Bij zieke planten wordt de fruitop-
Nog recenter werd Foc-TR4 waargenomen in Mo-
“Black Sigatoka” genoemd, wordt veroorzaakt
brengst gehalveerd. De schimmel infecteert alle
. Deze
door de schimmel Mycosphaerella fijiensis. Deze
banaan-, hoogland- en bakbanaanvariëteiten,
ziekte is duidelijk een nieuwe reële bedreiging voor
ziekte werd voor het eerst opgemerkt in de Si-
dus niet alleen de commerciële variant ‘Caven-
gatoka vallei in Fiji in 1912, maar was vermoe-
dish’. In Oost-Afrika daalde de bananenproduc-
planten in monoculturen
30, 31
.
en Indonesië tot de toplanden behoren wat bananenproductie betreft (zie Hoofdstuk 2). In oktober
2013 werd de uitbraak van deze stam in Jordanië
gerapporteerd32, waar relatief weinig bananen geteeld worden, ongeveer 40.000 ton per jaar, maar
32
zambique, Libanon, Oman en Pakistan
17, 18, 32
de volledige wereldproductie van bananen
.
33, 34
Banaan, het groene goud van het Zuiden
zit een bananenplant 11-16 bladeren, maar als
er door deze ziekte minder dan vijf overblijven,
zullen de vruchten zich niet voldoende kunnen
Figuur 3.3: Vernieling veroorzaakt door Sigatoka.
Er zijn weinig vruchten en gezond bladweefsel
te zien (Bron: J. Kimunye, IITA)
* Huidmondjes zijn structuren aan de oppervlakte van bladeren van landplanten. Een huidmondje bestaat uit een microscopisch kleine opening die
gevormd wordt door twee sluitcellen. Huidmondjes zijn zeer belangrijk
voor planten, omdat de plant via deze huidmondjes koolstofdioxide uit
de lucht opneemt en in het blad aangemaakte zuurstof en water de plant
weer afgeeft.
** Fotosynthese is een levensnoodzakelijk proces bij planten waarbij lichtenergie gebruikt wordt om koolstofdioxide om te zetten in energierijke
verbindingen (koolhydraten), zoals glucose. Op die manier is de plant in
staat zichzelf van energie te voorzien. De meeste fotosynthese vindt plaats
in de bladeren van een plant.
25
Virussen bedreigen de
bananenteelt wereldwijd
zen brengen het virus binnen in de vaatbundels van
en Costa Rica, maar recent ook in Ecuador49. Dit
genomen in de jaren 1960 in Ethiopië, bij een
de plant, waarna het zich vermenigvuldigt en zich
RNA-virus veroorzaakt spoelvormige paarsachtige
naaste variant van de banaan, namelijk de Ensete
over de volledige plant verspreidt. De eerste symp-
strepen op de bladeren, pseudostam, ter hoogte
ventricosum of de Ethiopische of valse banaan51.
tomen worden ongeveer 20 tot 25 dagen na infec-
van de hoofdnerven en zelfs op de vruchten. In-
In 2001 werd deze ziekte ook in Oeganda vastge-
(Bak)bananen worden bedreigd door meerdere
tie zichtbaar. Het virus veroorzaakt onregelmatige
fectie met dit virus, dat tussen bananenplanten
steld, waar het zich inmiddels over het volledige
virussen . Deze worden hoofdzakelijk via zuigen-
donkergroene vlekken en strepen van variabele
verspreid wordt door een bladluis, kan leiden tot
land verspreid heeft50. Ook andere Centraal- en
de insecten, zoals bladluizen, of door vegetatieve
lengte en grootte aan de bladschede en –stelen,
opbrengstverliezen van 30 tot 70%40.
Oost-Afrikaanse landen, zoals de Democratische
vermeerdering van de bananenplant verspreid.
alsook aan hoofd- en zijnerven
. De nieuwe jon-
Republiek Congo, Rwanda, Kenia, Tanzanië en
Vooral het ‘Banana bunchy top virus’ (BBTV) leidt
ge bladeren die zich aan een geïnfecteerde plant
Burundi zijn sindsdien getroffen22. De bacterie
tot zeer belangrijke economische verliezen.
ontwikkelen zijn veel smaller, korter, en vertonen
infecteert de meest geteelde bananenvariëteiten,
gele randen . Meestal wordt er geen of totaal mis-
maar de bierbanaan ‘Pisang Awak’ blijkt de gevoe-
BBTV is een klein virus met circulair enkelstrengig
vormd fruit geproduceerd. De ziekte werd voor het
ligste te zijn52, 53. Voornamelijk de mannelijke bloe-
DNA41. De verspreiding van plant tot plant gebeurt
eerst waargenomen bij de ‘Cavendish’ planten in
men zijn vatbaar voor infectie met Xanthomonas,
door middel van de bladluis Pentalonia nigronervo-
1889 in Fiji , terwijl de eerste infectie in 1901 op
want de plant wordt niet meer geïnfecteerd bij
sa . Tijdens het voeden verspreidt de bladluis het
het Afrikaanse continent, in Egypte, plaatsvond .
verwijdering van deze mannelijke bloemen.
virus van zieke naar gezonde planten. De bladlui-
Momenteel wordt BBTV-infectie bij bananenplan-
40
40, 41
43
42
44
45
ten waargenomen in 36 landen, waaronder 14 in
Wanneer een plant door de bacterie geïnfecteerd
. De verliezen
is, vergelen en verdorren de bladeren, rijpen de
zijn vaak zo drastisch (50 tot 100%), dat met de ba-
vruchten onregelmatig en onvolledig en vertoont
nanenteelt gestopt wordt.
het vruchtvlees gele en donkerbruine vlekken22.
Afrika en 22 in Azië en Oceanië
40, 46, 47
Bij doorsnede van de pseudostam, wordt een
Ook het ‘Banana streak virus’ (BSV) is wereldwijd
roze bacterieslijm waargenomen. De symptomen
verspreid en voor het eerst opgemerkt in 1958
verspreiden zich zeer snel doorheen de plant 3-4
in Ivoorkust . Dit virus bezit één dubbelstrengi-
weken na infectie en hangen af van de variëteit,
ge DNA-molecule en wordt naar andere planten
het groeistadium bij infectie en de manier van
overgedragen ofwel door de wolluis, ofwel door
overdracht. Bovendien blijken de symptomen zich
vermeerdering van besmette planten. Dit virus
vlugger te ontwikkelen tijdens vochtige seizoenen.
40
zorgt voor onregelmatige gele vlekken en strepen
Figuur 3.5: Symptomen van infectie met ‘Banana
De voornaamste bronnen van infectie zijn geïnfec-
op de bladeren en het vernietigen van de pseu-
Xanthomonas Wilt’ (Bron: V. Nakato, IITA)
teerde bananenplanten en resten ervan die op de
dostam door splitsing, wat meestal de dood van
grond blijven liggen, insecten, wind, regen, en ver-
de plant tot gevolg heeft. Dit virus infecteert ba-
vuild gereedschap. Wanneer de planten verwond
voor de eerste maal waargenomen in de Filipij-
De bacterie Xanthomonas
is de grootste vijand van
de banaan in Afrika
kan een besmet veld minstens 5-6 maanden niet
nen48, maar is nu ook al terug te vinden in een
De bananenziekte “Banana Xanthomonas Wilt”
gebruikt worden voor een nieuwe teelt22.
aantal andere Aziatische landen, waaronder Indië,
(BXW) is een ziekte die veroorzaakt wordt door
Figuur 3.4: Een bananenplant geïnfecteerd met het
Sri Lanka, Vietnam en Thaïland. In Latijns-Amerika
de bacterie Xanthomonas campestris cv. musacear-
BXW is een bedreiging voor de bananenpro-
‘Banana bunchy top virus’ (BBTV) (Bron: R.Swennen)
kwam de infectie voor het eerst voor in Colombië
um . Deze ziekte werd voor de eerste maal waar-
ductie en de bestaansmogelijkheden van kleine
nanenplanten in 43 landen, verspreid over Afrika,
Azië, Europa, Oceanië en tropisch Amerika.
De “Banana bract mosaic” ziekte werd pas in 1979
Banaan, het groene goud van het Zuiden
50
zijn door aaltjes of snuitkevers kunnen de bacteriën ook binnendringen via de wortels. Xanthomonas kan in afwezigheid van de bananenplant 3 tot
4 maanden in de bodem overleven54. Hierdoor
27
boeren in Centraal- en Oost-Afrika, aangezien de
en zo grote holtes vormen. Als gevolg kan de
overgebracht zijn naar New South Wales in 1910
volledige planten aangetast en vernietigd wor-
plant minder water en nutriënten vanuit de bo-
en naar Jamaïca in 191560. Er zijn aanwijzingen
den. Op die manier worden er geen vruchten
dem opnemen, waardoor de bladeren vergelen,
dat deze nematode voor 1960 niet aanwezig was
en dochterplanten/zijscheuten gevormd en gaat
vervroegd afvallen en de fruitproductie vermin-
in Afrika. Men vermoedt dat de sterke daling in
de plantage verloren53. De ziekte veroorzaakte al
dert en vertraagt. Door het verzwakte wortelge-
de bananenproductie sinds 1970 in Oeganda en
economische verliezen van 2-8 miljard dollar, en
stel valt de plant om
.
Tanzanië voornamelijk te wijten zijn aan de ne-
24, 56
matode Radopholus similis, hoewel deze nemato-
de verminderde productie zorgde ook voor enorme prijsstijgingen . Moko is een verwante ziekte
Nematoden hebben een levende gastheer nodig
de vaak samen met andere soorten voorkomt60.
die veroorzaakt wordt door de bacterie Ralstonia
om te overleven en zich voor te planten, maar
Radopholus similis bedreigt vooral de hoogland-
solanacearum. De oogstverliezen en symptomen
kunnen zich doorheen de bodem wel over een
bananen in Oeganda, maar nagenoeg alle bana-
zijn zeer vergelijkbaar met die van BXW. Moko
beperkte afstand van de ene naar de andere
nenvariëteiten zijn gevoelig voor infectie door
komt echter alleen voor in Latijns-Amerika .
gastheer verspreiden. De vrouwelijke nemato-
de andere soorten nematoden. Aangezien de
den leggen meerdere eitjes per keer in de ge-
infecties bij bananen vaak gepaard gaan met
Dwarse doorsnede van de pseudostam
Kleine wormpjes, grote problemen
ïnfecteerde wortels, en van ei tot ei duurt de le-
bijkomende besmettingen door schimmels en
(Bron: D. Coyne)
venscyclus van een nematode ongeveer 20 tot
bacteriën, is het moeilijk de impact van de nema-
Wereldwijd, maar vooral in Afrika, wordt de ba-
25 dagen . De jonge nematoden zullen ofwel in
todeninfectie te bepalen, maar de economische
met snuitkevers een plantage volledig vernielen.
nanenproductie zeer sterk bedreigd door infec-
de wortel blijven en deze verder infecteren ofwel
verliezen worden toch 30 tot 60% geschat20, 56.
Afgevallen plantenmateriaal op de grond vormt
tie met nematoden . De belangrijkste nema-
via de bodem naburige gezonde planten binnen-
toden zijn de migratorische Radopholus similis,
dringen. Verspreiding van nematodeninfecties
Pratylenchus goodeyi, Pratylenchus coffeae, Helico-
verloopt voornamelijk via besmette grond, bv. bij
tylenchus multicinctus en het wortelknobbelaaltje
overstromingen, besmet gereedschap, schoe-
Meloidogyne spp.
nen of het gebruik van ziek plantenmateriaal.
19
55
56
57-59
. Nematoden zijn microsco-
56
pisch kleine parasitaire wormen die zich voeden
Figuur 3.8: Schade veroorzaakt door snuitkevers.
dan ook een goede broedplaats voor de snuit-
De snuitkever,
een onderschatte vijand?
kevers. De snuitkevers vliegen zelden, maar ver-
De bananenboorder of snuitkever Cosmopolites
van de velden heel belangrijk om infecties vlug
spreiden zich al kruipend naar de verschillende
plantages61. Om verspreiding van een snuitkeverplaag tegen te gaan, is een goede bewerking
ten koste van planten. De bananenplant wordt
Infectie van bananenplanten met Radopholus si-
sordidus is een bedreiging voor bananenplanten
in te dijken en wordt best met ziektevrij planten-
geïnfecteerd via de wortels, waar de migratori-
milis werd voor het eerst waargenomen op Fiji in
in gebieden van Centraal-Afrika, Centraal-Ame-
materiaal gewerkt, bekomen na weefselcultuur.
sche nematoden zich met de celinhoud voeden
1891 . Geïnfecteerd plantenmateriaal zou dan
rika, Brazilië, Caraïben en Australië . De snuit-
Het is echter noodzakelijk om minstens twee
kever wordt geassocieerd met een snelle daling
jaar te wachten vooraleer een nieuwe plantage
van de opbrengsten in bananenplantages in
op voorheen besmette grond aan te planten61, 64.
60
61
Figuur 3.6: Bananenwortels geÏnfecteerd door Meloidogyne spp. Donkere won-
Figuur 3.7: Schade aangericht aan bananenplanten door
Oost-Afrika en met het “opbrengst daling syn-
den en necrotische wortels worden geïnduceerd door de voedende en gravende
nematodeninfectie (Bron: D. Coyne).
droom” in West-Afrika62. De snuitkever legt zijn
nematode (Bron: D. Coyne, IITA)
eieren aan de knol of de pseudostam van de
bananenplant. De groeiende larven graven tunnels, die tot 8 mm in diameter kunnen worden,
en de knol volledig doorzeven63. Vervolgens worden deze tunnels ingepalmd door schimmels en
verrot de knol. Sapstroom en opname van voedingsstoffen uit de bodem verminderen sterk,
de bladeren verwelken, enkel klein fruit wordt
gevormd en de planten vallen gemakkelijk om bij
windstormen61, 63. Op die manier kan een infectie
Banaan, het groene goud van het Zuiden
29
De boer als beschermheer van
zijn veld en bananenplanten
vlugger te herkennen en moesten op hun vel-
De meest vanzelfsprekende manier om versprei-
niet haalbaar vanwege te arbeidsintensief en te
ding van ziektes op het veld tegen te gaan is geïn-
duur66, 67. Verder lijkt de controle van de bana-
fecteerde planten zo snel mogelijk te verwijderen.
nenziektes, en vooral BXW, zeer sterk afhankelijk
Dit is echter niet altijd even eenvoudig. Verschil-
te zijn van land tot land en vooral van de poli-
lende ziektes, zoals de Panamaziekte, manifeste-
tieke stabiliteit. In Oeganda en Tanzanië, landen
ren zich voornamelijk eerst inwendig in de plant
met sterke politieke leiders en stabiliteit, is BXW
en worden pas zichtbaar wanneer een algemene
voor meer dan 90% onder controle. In de oos-
aantasting van meerdere planten al een feit is.
telijke regio’s van de Democratische Republiek
Verder kunnen schimmelsporen zich zeer vlug
Congo is de ziekte echter verviervoudigd.
den proper gesteriliseerd materiaal gebruiken.
Hierdoor werd de ziekte tot 10% gereduceerd.
De maatregelen bleken voor de boeren echter
verspreiden, waardoor de ziekteuitbreiding zodanig snel is dat bij de tijd dat de boer reageert het
Om de verspreiding van plagen en ziektes te-
al te laat kan zijn. De enige oplossingen zijn de
gen te gaan is het zeer belangrijk dat de boeren
plantage volledig te vernietigen, alle wortels uit de
steeds met ziektevrij plantenmateriaal werken.
bodem te verwijderen en de gronden een aantal
maanden braak te laten liggen of te beplanten
4
Biotechnologie kan de banaan
beschermen tegen zijn vijanden
Verschillende ziektes, veroorzaakt door bacteriën, schimmels, virussen, nematoden en
insecten, bedreigen de wereldwijde productie van bananen. Dit is niet alleen een groot
probleem voor de westerse wereld die de bananen voornamelijk importeert, maar vooral
voor de kleine, lokale boeren in ontwikkelingslanden die bananen telen als bron van voedsel
en inkomen. Om deze ziektes te bestrijden kunnen verschillende pistes bewandeld worden.
Methodes zoals preventie en/of beter teeltmanagement lijken eenvoudig, maar zijn niet
altijd even efficiënt. Sproeien met fungiciden en/of pesticiden, als deze al beschikbaar zijn,
kunnen, vooral bij veelvuldig gebruik, een negatieve impact hebben op mens en milieu. Bij
veelvuldig gebruik kan de doeltreffendheid bovendien sterk verminderen door mogelijke
resistentie van de ziekteverwekkers. Via veredeling wordt getracht om voor elke plaag
ziekteresistente bananensoorten te genereren, wat eenvoudiger gezegd dan gedaan blijkt.
Daarom is men ervan overtuigd dat het bekomen van resistente bananenplanten door
middel van biotechnologie een verantwoorde methode is die doeltreffende oplossingen
kan bieden.
met andere, niet-gevoelige planten, totdat alle
verschaffen over de potentiële impact en vroege
WEEFSELCULTUUR OM ZIEKTEVRIJ
PLANTENMATERIAAL TE ONTWIKKELEN
opsporing van infectiesymptomen65.
Weefselkweek of weefselcultuur is een methode om planten
ziekteverwekkers verdwenen zijn29. Daarom is
het belangrijk de telers voldoende informatie te
te vermeerderen onder steriele condities en is gebaseerd op
Om plagen te vermijden of in te perken is het
het feit dat individuele plantencellen in staat zijn om tot een
ook van belang de planten ver genoeg uit el-
volwassen plant (totipotentie) uit te groeien. Weefselcultuur
kaar te planten, voor een betere waterdrainage
is heel belangrijk voor verscheidene bananensoorten wegens
en onkruidbestrijding te zorgen, de mannelijke
hun nagenoeg steriele karakter en de moeilijkheid zich voort te
knoppen te verwijderen, en met ontsmet ge-
planten via zaad. Door middel van weefselcultuur of vegetatie-
reedschap te werken. Dit blijkt echter niet haal-
ve vermeerdering kan de plant zich op een eenvoudige manier
baar voor de lokale boeren omdat dit extra werk
vermeerderen en kunnen zo genetisch identieke nakomelingen
en kosten met zich meebrengt66. In 2001 werd in
(klonen) gegenereerd worden. Zodra nieuwe groeischeuten
Oeganda een strategie ontwikkeld om BXW zo-
zich ontwikkelen, kunnen ze van de plant afgesneden worden
veel mogelijk in te dijken: alle geïnfecteerde plan-
en na behandeling in een steriele omgeving met gefilterde lucht
ten dienden verwijderd en verbrand te worden,
opgegroeid worden op een vaste voedingsbodem, die zouten,
de bananenplanten mochten slechts beperkt
nutriënten, vitamines en plantenhormonen bevatten voor op-
vervoerd en verplaatst worden en de mannelijke
timale plantenontwikkeling. Door het toedienen van het plan-
vruchtknoppen werden weggenomen. De boe-
tenhormoon auxine aan de voedingsbodem zal de scheut wor-
ren kregen bijkomende informatie om de ziekte
tels vormen. Eenmaal de scheut groot genoeg is, kan deze in
aarde geplant worden.
(Bron: Sebastien Carpentier, KU Leuven) Banaan, het groene goud van het Zuiden
31
Wie van de drie: monocultuur,
mengcultuur of gewasrotatie?
‘Gros Michel’ op de grote plantages niet meer
zijn, maar door de sterk verhoogde opbrengst
Een belangrijke oorzaak voor de snelle versprei-
boeren, die bananen en koffiestruiken samen te-
sity International samen met nationale program-
zagen de boeren hun inkomen met maar liefst
ding van ziektes bij de huidige (commerciële)
len, waardoor ze minder last hebben van infectie
ma’s in Afrika om zoveel mogelijk geïnfecteerde
145% stijgen . Het gebruik van scheuten uit
bananen is dat op de plantages voornamelijk
met Fusarium oxysporum72. Bananen worden ook
planten van de velden te verwijderen en in gese-
weefselcultuur verschaft ook toegang tot nuttige
aan monocultuur gedaan wordt30. Monocultuur
dikwijls samen geteeld met bonen (Phaseolus
lecteerde gebieden nieuw gezond plantenmate-
endofyten in de bodem. Endofyten zijn nuttige
betekent dat op hetzelfde stuk grond altijd het-
vulgaris)73. Voor de teelt van bonen is er echter
riaal aan te planten (www.rtb.cgiar.com). Het ver-
micro-organismen, zoals bacteriën en schimmels,
zelfde gewas geteeld wordt en er dus niet aan
meer grondbewerking nodig, wat nadelig kan zijn
krijgen van ziektevrij plantenmateriaal is namelijk
die samenleven met de bananenplant en de plant
vruchtwisseling gedaan wordt. Bij banaan is dit
voor de bananenplanten wegens eventuele be-
niet zo eenvoudig. Voorlopig is het International
beschermen tegen ziektes, plagen en ongunstige
nog een extreme vorm van monocultuur, aange-
schadiging van de wortels74.
Transit Centre gehuisvest in het Laboratorium
condities, zoals droogte en warmte. Een belang-
zien de jonge bananenplantjes allemaal genetisch
voor Tropische Plantenteelt (KU Leuven – Rony
rijke kanttekening is wel dat bananenplantjes,
identiek zijn aan de moederplant, met alle goede,
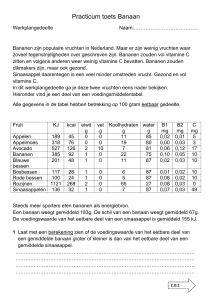
Een efficiënte manier om ziektenverspreiding
Swennen, zie kaderstuk Hoofdstuk 2) het belang-
bekomen na weefselcultuur, iets gevoeliger blij-
maar ook alle slechte/negatieve kenmerken.
tegen te gaan is wisselbouw of gewasrotatie.
rijkste om 100% ziektevrij kiemmateriaal te beko-
ken te zijn voor de Panamaziekte dan op het veld
men na weefselcultuur.
ontstane scheuten29, 71. Toch wordt de industrie
Vooral op plantages waarop exportbananen ge-
gewassen na elkaar geteeld. Pas na enkele ja-
omtrent weefselcultuur steeds belangrijker voor
teeld worden, komt enkel de genetisch identieke
ren komt hetzelfde gewas terug op het perceel.
Weefselcultuur bevordert de verspreiding van
de productie van bananen in Afrika. Een aantal
‘Cavendish’-banaan voor. Wanneer deze planten
Deze wisselbouw zorgt daarenboven ook voor
ziektevrij plantenmateriaal en levert talrijke voor-
KMO’s en onderzoekscentra specialiseerden zich
gevoelig zijn voor een bepaalde schimmel, virus,
een betere bodemstructuur en –vruchtbaarheid
delen op voor de lokale boeren68-70. Belangrijk
in deze techniek om zo tegemoet te komen aan
bacterie en/of nematode die zich in de bodem
en doorbreekt elk jaar de samenstelling van de
hierbij is wel dat dit materiaal enkel uitgeplant
de steeds groeiende vraag van de boeren.
bevindt, gebeurt de verspreiding naar alle plan-
bodem, zowel naar populatie van micro-organis-
ten razendsnel. De kleine boeren zorgen echter
men als chemische eigenschappen. Recent kon
steeds meer voor teeltwisseling op hun land,
aangetoond worden dat het afwisselend telen
omdat ze beseffen dat telen van bananenvarië-
van ananas en banaan effectiever is in de be-
teiten met verschillende gevoeligheid voor een
strijding van de Fusarium-populatie in de bodem
bepaalde plaag de verspreiding ervan vertraagt
dan van banaan en maïs75. Weinig boeren in het
en hiermee de kans op een geslaagde oogst ver-
Zuiden zijn echter geneigd om aan gewasrotatie
hoogt. Daarom worden meestal 4-5 verschillen-
te doen, omdat de bananenplant een meerjarig
de bananenvariëteiten gegroeid, maar dit aantal
gewas is met een lange cyclusduur en zeer be-
kan zelfs oplopen tot 20.
langrijk is voor hun voedsel en inkomen.
Een stapje verder in de goede richting is het om-
manier wordt aan mengcultuur gedaan en kun-
Zijn chemische, biologische of
fysische bestrijdingsmiddelen
de redders voor de banaan?
nen de ziektes zich minder snel verspreiden.
Zowel voor het bestrijden van de Panamaziekte
Figuur 4.1: Mengcultuur van bananen en bonen
Voor export werd de ‘Gros Michel’ volledig ver-
als Black Sigatoka werden verschillende schim-
in Tanzanië (bron: R. Swennen)
vangen door de ‘Cavendish’, omdat de teelt van
melbestrijdende middelen of fungiciden uitge-
De kleine boeren gebruiken echter vooral jonge
bruik te maken van uit weefselcultuur bekomen
scheuten afkomstig van oude plantages om de
plantjes, verhoogden de bananenboeren in Kenia
kosten te drukken, waardoor de verspreiding
hun opbrengst van 10 naar 30 ton per hectare .
van de bananenziektes alleen nog in de hand ge-
De aankoop van de plantjes bleek wel duur te
werkt wordt. Momenteel werken IITA en Biover-
68
68
wordt op niet-geïnfecteerde gronden. Door ge-
se landen, zoals Costa Rica en Nicaragua, wordt
‘Gros Michel’ echter nog wel geteeld door kleine
Hierbij worden op een stuk grond verschillende
geven van bananenplanten door andere planten, zoals koffie, sorghum, aardappelen, maïs,
cassave, rijst, suikerriet of jamwortel24. Op die
Banaan, het groene goud van het Zuiden
mogelijk is. In vele Zuid-en Centraal-Amerikaan-
33
test36, 76. De fungiciden tegen Fusarium oxysporum
kel op commerciële bananenplantages gebruikt,
bleken goed in vitro (bv. in een proefbuis) en in
waar de bananenopbrengst dan wel met 250%
de plantenkamer te werken, maar niet effectief
kan stijgen. Nematiciden verdelgen de nemato-
te zijn op het veld. Zelfs injectie van de bana-
den in de bodem niet, waardoor de behandeling
nenplanten met of het dippen van de planten in
regelmatig herhaald moet worden en de kans op
fungiciden leverde geen overtuigende resultaten
resistente stijgt.
op
77, 78
Een werk van lange adem: het
ontwikkelen van resistente bananen tegen ziekten en plagen
via kruisingsprogramma’s
. Grondontsmetting, warmte sterilisatie of
gens de relatief lange generatietijd, zal de gewenste hybride pas na een groot aantal jaren bekomen
worden. (4) Een bananenplant is eerder groot, dus
er is zeer veel plaats nodig om alle bekomen hybriden uitvoerig te analyseren. (5) Het genoom is nog
onvoldoende gekend om alle genen die nodig zijn
om resistentie tegen een bepaalde plaag te iden-
grondoverspoeling leken kortstondig het aantal
Om virale infecties bij bananenplanten zoveel
Niettegenstaande (bak)bananen wereldwijd zeer
tificeren4, 89. Desondanks werden reeds een aantal
schimmelbesmettingen te reduceren, maar heel
mogelijk te bestrijden wordt het herbicide 2,4D
belangrijke voedingsgewassen zijn, zijn tot op
successen genoteerd4, 85, 89, 94.
snel werd de grond opnieuw met de schimmel
gebruikt en dit in combinatie met een insecticide
vandaag slechts zeven veredelingsprogramma’s
die zoveel mogelijk de bladluis, de drager van het
opgesteld met als doel betere bananensoorten
De eerste bananenhybriden werden ontwik-
geïnfecteerd
29, 79
.
virus, vernietigt. Deze behandelingen kunnen
te ontwikkelen
. Deze programma’s komen
keld in Trinidad, maar dat programma bestaat
Jaarlijks zijn 50 tot 70 sproeibeurten met fungici-
echter wegens de dure kostprijs niet door alle
voor in Brazilië, Kameroen, Ivoorkust, Guade-
niet meer. Later maakte Fundación Hondureña
den vereist om Black Sigatoka te bestrijden, wat
boeren toegepast worden.
loupe, Honduras, Indië, Nigerië, Tanzanië en
de Investigación Agrícila (FHIA) in Honduras
Oeganda . Ze hebben niet alleen tot doel plan-
(www.fhia.org.hn) nieuwe hybriden. Hoewel de
socio-economisch en ecologisch onverantwoord
4, 89
4
. De kosten lopen vlug op tot ongeveer 1800
Voor de bestrijding van de Xanthomonas bacterie
ten met verhoogde resistentie tegen ziekten en
initiële focus op dessertbananen lag, wordt nu
dollar per jaar, wat teveel is voor de kleine loka-
zijn momenteel geen pesticiden of biologische
plagen te ontwikkelen, maar spitsen zich ook toe
ook met kook- en bakbananen gewerkt. Om-
. Bij grote plantages worden deze
agentia beschikbaar . Ook voor de andere ba-
op het genereren van planten met verhoogde
dat vruchtbare planten nodig zijn voor de ver-
fungiciden meestal toegediend met vliegtuigen
nanenziekten lijken biologische bestrijdingsmid-
opbrengsten, resistenties tegen extreme omge-
edeling, gebruikt FHIA als moederplant heel
of helikopters, waardoor ook extra landings-
delen weinig effectief . Bij biologische bestrij-
vingsomstandigheden, zoals droogte, en op het
dikwijls ‘Highgate’ (AAA), een mutant van ‘Gros
plaatsen en brandstof nodig zijn. Aldus kunnen
ding van een plaag worden micro-organismen of
reduceren van oogstverliezen door vroege rij-
Michel’ (AAA). Deze plant geeft na bevruchting
de bestrijdingskosten oplopen tot 20-40% van
biologische grondstoffen toegevoegd aan de bo-
ping geïnduceerd door transportschade of door
immers aanleiding tot een aantal vruchten met
. Het
dem om de aanvallers te lijf te gaan. Biologische
insecten . Bij de meeste kruisingsprogramma’s
zaden. Ook AAB soorten, zoals ‘Dwarf Brazili-
veelvuldig gebruik van fungiciden zorgt er ook
controle van de Panamaziekte lijkt momenteel
ligt de nadruk op het ontwikkelen van hybriden
an/Dwarf Apple’ worden frequent benut. Eén
voor dat de schimmels steeds toleranter en re-
nog onmogelijk. Toch blijken een aantal bodems
(nakomelingen afkomstig uit twee verschillende
van de eerste hybriden die in 1988 ontwikkeld
sistenter worden tegen deze bestrijdingsmidde-
te bestaan die de verspreiding ervan kunnen
oudersoorten) die resistentie vertonen tegen de
werd was FHIA-01 of ‘Goldfinger’, een tetraplo-
len, met het gebruik van steeds giftigere fungici-
onderdrukken, maar het is tot nu toe echter
bladvlekkenziekte, maar ook tegen nematoden
ïde dessertbanaan die resistentie vertoont te-
den of combinaties van fungiciden tot gevolg .
onduidelijk welke bodemfactoren daarvoor ver-
en Fusarium .
gen de bladvlekkenziekte, Fusarium FocTR 1 en
is
37, 80
le boeren
16, 81
de totale productiekost van de banaan
36, 80
36
22
29
90
91
antwoordelijk zijn85. Voor het verdelgen van de
Foc-TR4, alsook verhoogde resistentie tegen de
Nematoden worden voornamelijk bestreden
nematode Radopholus similis in de bananenplant
De kruisingsprogramma’s voor banaan kennen
nematode Radopholus similis, maar niet tegen
met chemische bodemontsmettingsmiddelen
‘Grand Nain’ kon recent aangetoond worden
minstens vijf belangrijke belemmeringen. (1) De
andere nematoden. Daarnaast blijkt deze hy-
en nematiciden , die echter niet doeltreffend
dat het vloeibare bijproduct van de productie
meest frequent geconsumeerde bananen zijn
bride ook tolerant te zijn tegen koude en wind
zijn en voor veel problemen zorgen, zoals ver-
van sisalvezels heel efficiënt werkt86. Of dit een
zaadloos, steriel en triploïd. Daarom moeten di-
(www.fhia.org.hn). Aanvankelijk leek deze hy-
vuiling van bodem en grondwater en gezond-
algemeen bestrijdingsmiddel tegen nematoden-
ploïde, vruchtbare, ouders gebruikt worden die
bride ook resistent te zijn tegen de “Black Leaf
. Tot vóór
infecties voor alle bananensoorten zal worden
echter meestal zwakke agronomische kenmerken
Streak” schimmelinfectie, maar deze resistentie
enkele jaren werd methylbromide frequent ge-
valt nog af te wachten. Ook de aanwezigheid van
hebben en waarvan de vruchten dikwijls niet van
werd al vlug door de schimmel doorbroken4. De
bruikt om nematodenpopulaties te doden, maar
algen of een extract van Camellia-zaden induce-
goede kwaliteit zijn. (2) Het kruisen van bananen
FHIA-01 hybride heeft echter een lagere vrucht-
het gebruik ervan wordt momenteel beperkt
ren een sterke daling in de nematodenpopulatie
vergt zeer veel handenarbeid (zie verder). (3) Het
vlees/schil verhouding en is iets minder lekker
door zijn negatief effect op de ozonlaag. Door
in de bodem en hebben een positieve invloed op
inkruisen van één of meerdere genen verloopt
en daardoor minder geliefd dan de ‘Cavendish’
de hoge kostprijs worden nematiciden ook en-
de groei van de bananenplant
en, we-
soorten ‘Grand Nain’ en ‘Williams’. Deze hybride
82
heidsproblemen bij mens en dier
Banaan, het groene goud van het Zuiden
83, 84
87, 88
.
steeds over verschillende generaties
92, 93
35
wordt daarom voornamelijk door kleine boeren
NARITA (NARO-IITA) hybriden op, waarvan er nu
domestication” noemt99. Bij “superdomestication”
dat de bloemenbundel gedurende ongeveer
en niet als exportproduct geteeld. De hybride
25 getest worden in Oeganda en Tanzanië op
is er een sterke interactie tussen telers en weten-
één week elke dag manueel bestoven wordt. De
FHIA-03, de ’Sweetheart’ kookbanaan, is een te-
het veld . Deze hybriden zijn allemaal afkomstig
schappers met genoomkennis, die samen op pa-
meeste bestoven vrouwelijke bloemen zullen
traploïde banaan die geen zaden produceert en
van een kruising tussen verschillende vrouwelij-
pier het profiel van de hybride met alle gewenste
geen zaad produceren, sommigen een paar en
een karakteristieke lichtgroene pseudostam be-
ke triploïde vruchtbare EAHB-hooglandbananen,
eigenschappen opmaken, waarna kan bepaald
een beperkt aantal veel zaden. Het grote pro-
zit. FHIA-03 vertoont resistentie tegen Fusarium,
verschillende diploïden en de diploïde Calcutta 4
worden hoe die nieuwe hybride zal ontwikkeld
bleem is dat deze zaden meestal niet vanzelf kie-
de bladvlekkenziekte en gedeeltelijke resistentie
banaan (AA) die zeer vruchtbaar pollen bezit en
worden en welke specifieke genen noodzakelijk
men en daarbij hulp nodig hebben van de ver-
tegen de nematode Radopholus similis (www.
resistentie vertoont tegen de bladvlekkenziekte .
zijn om het gewenste resultaat te bereiken .
edelaars. De veredelaars verzamelen daarom de
fhia.org.hn). Deze hybride wordt commercieel
De geselecteerde tetraploïden werden vervol-
geteeld in Cuba op meer dan 3500 hectaren,
gens met verbeterde diploïden gekruist om zo
zonder veelvuldig toedienen van meststoffen en
een triploïde hybride te bekomen.
96
4
fungiciden . Verder wordt deze hybride ook ge85
90
embryo’s van de gesteriliseerde zaden en laten
Monnikenwerk: Veredelen van
bananen in de praktijk
deze vervolgens ontkiemen in proefbuizen met
vast medium die alle nodige voedingsstoffen
bevatten. Nadien worden deze plantjes overge-
teeld in Burundi, Kameroen en Nigerië. De FHIA-
Ondanks de eerste successen staan de veredeling-
17, FHIA-23 en FHIA-25 hybriden worden succes-
programmas’s nog voor een aantal grote uitdagin-
Hoewel de meeste bananensoorten die vandaag
“afharden” om dan uiteindelijk in het veld geplant
vol geteeld in de Kagera streek van Tanzanië na
gen. Voor de consument is het vooral belangrijk
geteeld en gegeten worden triploïd, steriel en
te worden. In de meeste gevallen worden de tri-
verdeling van 4 miljoen scheuten. Hierdoor zijn
dat de hybride bananen dezelfde kwaliteit (smaak,
zaadloos zijn, zijn er toch een aantal soorten die
ploïde nakomelingen geëvalueerd als potentieel
de inkomens van 0,5 miljoen mensen met een
geur, afwezigheid van zaden) bezitten als de hui-
een klein beetje vruchtbaarheid blijven behouden
nieuwe variëteiten en de diploïde (2 sets van
factor 3 verhoogd.
dige niet-resistente bananen. Verder bestaat het
en zaad kunnen vormen wanneer ze bestoven
chromosomen) en tetraploïde (4 sets van chro-
gevaar dat de ziektenverwekkers, zoals schimmels,
worden met sterk levensvatbaar pollen. De ver-
mosomen) planten als nieuwe kandidaat-ouders
IITA (Nigerië) startte zijn veredelingsprogram-
nematoden en virussen, de resistentie doorbre-
edelaar speelt hier de rol van natuurlijke bestuiver.
voor volgende kruisingen. Zulke kruisingen uit-
ma’s voor banaan in de jaren 1980 toen de blad-
ken, vooral als de resistentie slechts door één gen
De veredelaar verzamelt ’s morgens de manne-
voeren bij bananenplanten is duidelijk een zeer
vlekkenziekte voor het eerst in die regio opge-
bepaald wordt . Daarnaast verleent een gen soms
lijke bloemen en brengt ze naar de vrouwelijke
ingewikkelde techniek die de nodige vakman-
merkt werd. Al snel werd vooruitgang geboekt
resistentie tegen één type nematode, bv. Radopho-
bloemen van de te bevruchten plant. Iedere dag
schap en uitgeruste apparatuur vereist die niet
door vruchtbare bakbanaansoorten te kruisen
lus similis, maar daarom niet tegen andere belang-
openen de bloemen zich achtereenvolgens, zo-
beschikbaar is voor de kleine boer.
met wilde resistente voorouders95. In 1995 start-
rijke nematoden, zoals Pratylenchus spp.89, 98 Dit
te IITA samen met de “National Agricultural Re-
impliceert dat verschillende resistentiegenen in de
search Organization” (NARO) een kruisingspro-
hybride dienen binnengebracht te worden, wat het
gramma in Oeganda omdat de ziekte ook daar
kruisingsprogramma complexer en langer maakt.
was uitgebroken, met bijzondere aandacht voor
Tenslotte zijn er nog geen bananen gekend die
de Oost-Afrikaanse hooglandbananen. Vrucht-
resistent zijn voor de bacterie Xanthomonas noch
bare EAHB-soorten werden geïdentificeerd, re-
voor het ‘Banana bunchy top’ virus (BBTV).
97
plant naar aarde, laat men ze verder groeien en
sistente tetraploïde ouders werden ontwikkeld
en kruisingen werden uitgevoerd om resistente
Voor de veredelingsprogramma’s is het vooral
triploïde hybriden te bekomen. In 2010 bracht
belangrijk dat in de toekomst de genomen van
NARO “Kabana 6” (Kiwangaaazi) op de markt.
de verschillende bananensoorten nog meer in
Deze hybride geeft een hoge opbrengst en ver-
detail bestudeerd worden en de genen met hun
toont resistentie tegen de bladvlekkenziekte en
bijhorende eigenschappen nog beter in kaart ge-
gedeeltelijke resistentie tegen nematoden en
bracht worden. Deze genomische kennis kan dan
snuitkevers . De samenwerking tussen NARO
leiden tot een vluggere ontwikkeling van de ver-
en IITA leverde ondertussen 27 verschillende
edelingsprocedures, een proces dat men “super-
89
Figuur 4.2: Manueel bevruchten van
Banaan, het groene goud van het Zuiden
bananenplanten (Bron: M. Batte)
37
VEREDELING VERSUS GENETISCH GEWIJZIGDE PLANTEN100
Met gentechnologie en het genetisch wijzigen van planten wil men hetzelfde doel bereiken als bij klassieke verede-
Al van bij het ontstaan van de landbouw gaan mensen op zoek naar betere, sterkere, en commercieel interessante
voordeel van gentechnologie is dat men heel gericht één of meerdere gewenste eigenschappen kan binnenbrengen
planten. Deze nieuwe en verbeterde planten worden verkregen door bij elke oogst continu te selecteren. Door twee
in een plant. De genetisch gewijzigde (GGO) plant zal hierdoor genetisch identiek zijn aan de interessante ouder-
verschillende ouderplanten met elkaar te kruisen, worden hun eigenschappen in één plant verenigd. Vervolgens
plant 2 (zie figuur 4.3), behalve voor het kenmerk dat werd ingevoerd. Hierdoor wordt de nieuwe interessante plant
wordt enkel verder gewerkt met de nakomelingen die de interessante combinatie bezitten van de eigenschappen
veel vlugger bekomen dan bij klassieke veredeling. Een ander groot voordeel van gentechnologie is dat het nieuwe
van de twee ouderplanten. Oorspronkelijk gebeurden deze kruisingen en selecties zonder te beseffen wat op het
ingevoerde DNA niet alleen afkomstig kan zijn van dezelfde plantensoort, maar ook van andere plantensoorten,
niveau van het erfelijk materiaal, het DNA, gebeurde. Nu weet men dat tijdens het kruisen de planten grote stukken
en zelfs van gisten, dieren en schimmels. Met andere woorden, de soort grenzen bestaan niet meer.
ling, namelijk nieuwe rassen creëren met interessante eigenschappen voor boer, industrie of consument. Het grote
DNA uitwisselen, waarbij de helft van het DNA van ouderplant 1 wordt verenigd met de helft van het DNA van
ouderplant 2 (zie figuur 4.3). Zo ontstaan nakomelingen die elk een unieke combinatie bevatten van de gewenste
gewenst kenmerk van ouderplant 1 overbrengen naar ouderplant 2. Hiervoor zijn verschillende opeenvolgende
GENETISCH WIJZIGEN VAN PLANTEN DOOR MIDDEL VAN HET
GENENKANON OF VAN AGROBACTERIUM TUMEFACIENS1 101
kruisingen nodig, waarbij de nakomelingen met de gewenste eigenschap steeds opnieuw gekruist wordt met de
Planten kunnen via verschillende manieren genetisch gewijzigd worden, maar tot op vandaag zijn de meest ge-
oorspronkelijk (commercieel interessante) ouderplant 2. Dit proces noemt men terugkruisen. Het terugkruisen
bruikte methoden het genenkanon of Agrobacterium tumefaciens. DNA binnenbrengen in de plant via het ge-
wordt verschillende malen herhaald zodat een nieuwe plant bekomen wordt die zoveel mogelijk eigenschappen
nenkanon is een mechanische manier, die ook wel “particle bombardment” genoemd wordt. Minuscuul kleine
bezit van de oorspronkelijk interessante ouderplant 2, maar met de nieuwe eigenschap van ouderplant 1. Afhan-
goudbolletjes worden bedekt met het DNA dat men in de plant wil binnenbrengen. Die goudbolletjes worden
kelijk van de duur van de levenscyclus (van zaad tot zaad) en de voorplantingsmethode van de plant kan deze
vervolgens onder hoge druk in het plantenweefsel “geschoten” en in sommige gevallen dringt het DNA binnen in
procedure verschillende jaren tot verschillende tientallen jaren in beslag nemen. Voor gewassen die voornamelijk
de kern van de plantencel, waar het spontaan ingebouwd wordt in het DNA van de plant. Deze methode is echter
vegetatief voortplanten, zoals aardappel, zoete aardappel, banaan en cassave, zijn zeven generaties nodig, voor
niet zo efficiënt en dikwijls worden veel kopijen en/of slechts stukken van het gewenste DNA in het planten-DNA
zelf-bestuivende gewassen, zoals rijst en tarwe, 10 generaties, en voor kruisbestuivende gewassen, zoals maïs,
ingebouwd. Een veel efficiëntere transformatiemethode maakt gebruik van het natuurlijk DNA-overdrachtsmecha-
zelfs 17 generaties . Bij deze veredeling kunnen dus enkel eigenschappen gecombineerd worden van één planten-
nisme van de bodembacterie Agrobacterium tumefaciens.
en ongewenste eigenschappen van de ouderplanten. Gedurende eeuwen selecteerden de landbouwers op deze
manier de minder interessante eigenschappen weg. In sommige veredelingsprogramma’s wil men vaak maar één
93
soort of van nauw verwante soorten (bv. banaan met banaan, maar niet met tomaat of rijst).
Figuur 4.4: Genetische transformatie met behulp
van Agrobacterium tumefaciens
Figure 4.3: Conventionele veredeling versus biotechnologie
Conventionele veredeling
eigenschap uit plant 1 die men in plant 2 wil inbrengen
CHOMOSOMAAL DNA
CHROMOSOOM
T - DNA
TI PLASMIDE
8 tot 10 kruisingen met plant 2
Biotechnologie
AGROBACTERIUM TUMEFACIENS
T - DNA
PLANTENCEL
In de natuur infecteert deze bacterie bepaalde plantencellen en brengt tijdens dat infectieproces een stukje van
zijn eigen DNA, het transfer of T-DNA, over naar de kern van de plantencel waar het planten-DNA zich bevindt.
Dit T-DNA wordt in het planten-DNA ingebouwd. Doordat dit T-DNA de informatie draagt voor de aanmaak van
opines, moleculen waarmee Agrobacterium zich voedt, verplicht de bacterie de plant om zijn voedsel te produ-
Banaan, het groene goud van het Zuiden
39
ceren. In de jaren 1970 ontrafelden moleculair-biologen, waaronder de Gentse professoren Marc Van Montagu
en Jeff Schell, het mechanisme van DNA-overdracht. Ze kwamen tot de ontdekking dat de genen die op het T-DNA
liggen niet essentieel zijn voor de overdracht van het bacterieel DNA naar de plantencel. Ze konden dit DNA vervangen door het gewenste DNA en de transfer naar de plantencel bleef doorgaan. Op die manier maakte de plant
geen opines meer aan voor de bacterie, maar bracht de bacterie voor de mens interessante informatie over naar
de plant.
Recent werd aangetoond dat alle 291 geteste gecultiveerde zoete aardappelvariëteiten 1 of meerdere T-DNA genen
bezitten, wat impliceert dat deze transfer van T-DNA-genen reeds lang geleden tijdens de evolutie plaatsgreep102.
Naast het gewenste DNA kan ook steeds het DNA van een selectiegen ingebouwd worden tijdens de genetische
transformatie. Genetische transformatie van cellen gebeurt immers niet met 100% efficiëntie en de planten die
de nieuwe genetische informatie kregen moeten opgespoord worden. Het selectiegen zorgt ervoor dat cellen die
het nieuwe DNA opgenomen hebben weerstand bieden tegen een bepaald herbicide of antibioticum, waardoor
alle plantjes na genetische modificatie gewoon opgegroeid kunnen worden in aanwezigheid van dat bepaalde
herbicide of antibioticum. Enkel de plantjes die overleven hebben het nieuwe DNA ingebouwd gekregen en de rest
sterft af. Deze selectie gebeurt tijdens de ontwikkelingsfase in het laboratorium en het antibioticum wordt in het
veld niet gebruikt.
Figuur 4.6: Verschillende stadia tijdens het transformatieproces. (A, B) ECS-cellen, (C) Regeneratie naar callus, (D, E) Ontwikkelen
van GGO-scheuten, (F) GGO-planten zijn overgeplant naar aarde (Bron: R. Swennen en B. Panis)
Voor beide methoden, het genenkanon en de transformatie met Agrobacterium tumefaciens, wordt van de
unieke eigenschap gebruik gemaakt dat individuele plantencellen in staat zijn om uit te groeien tot een volledig
nieuwe plant. Dit betekent dat uit één GGO-plantencel een volledige plant kan bekomen worden waarvan elke cel
het nieuwe DNA bezit.
Genetische transformatie
van banaan: de praktijk
hun eigen DNA en anderzijds kunnen uitgroeien
tot een volledige plant. Voor bananentransformatie worden daarom embryogene celsuspensie (ECS)-cellen gebruikt103, 109, 110.
Sinds 20 jaar is het mogelijk om bananen genetisch te wijzigen door het inbrengen van vreemde
De eerste en tevens moeilijkste stap in het trans-
genen, ofwel met behulp van het genenkanon103,
formatieproces is de ontwikkeling en isolatie van
, ofwel door middel van Agrobacterium tume-
deze ECS-cellen, die uit meristemen, embryo’s of
104
faciens (zie kaderstuk)
. Hoewel de transfor-
onrijpe vrouwelijke bloemen kunnen bekomen
matieprocedure voor elke bananensoort lichtjes
worden80, 111. De aanmaak van een suspensie kan
kan variëren, is ze toch steeds volgens hetzelfde
tot 18 maanden duren. Deze ECS-cellen worden
principe opgebouwd en gebeurt ze in een volle-
6 tot 7 dagen in een Agrobacterium-suspensie
dig steriele omgeving.
geïncubeerd, waarna ze gewassen worden en de
105-109
bacteriën gedood worden. De cellen worden ver-
Banaan, het groene goud van het Zuiden
Bij deze transformatiemethode is het heel be-
volgens op medium met een antibioticum gelegd
langrijk “transformatie-competent weefsel” te
en krijgen gedurende 2 tot 3 maanden de tijd om
isoleren, namelijk cellen die enerzijds in staat zijn
uit te groeien tot een GGO-plant. De ECS-cellen
om vreemd DNA op te nemen en in te bouwen in
die het nieuwe DNA niet opgenomen hebben
41
zullen reeds vroeg afsterven. Nadien worden de
en/of in de plantenkamer en pas daarna in het
GGO-planten overgezet naar aarde waar ze kun-
veld. De redenen hiervoor zijn dat (1) screening
nen uitgroeien tot een volwassen plant112. Ver-
in vitro vlugger gaat, (2) de optimale groeicon-
mits de GGO-plant uit één getransformeerde cel
dities voor bananen in vitro en in groeikamers
ontwikkeld is, bevatten alle cellen in die GGO-ba-
kunnen gesimuleerd worden (hoge vochtigheid
nanenplant hetzelfde nieuwe DNA op dezelfde
en hoge temperatuur), en (3) hiermee plaats be-
plaats106. De volledige procedure neemt ongeveer
spaard wordt, daar bananenplanten tot wel 4
6 tot 8 maanden in beslag en is heel reproduceer-
meter hoog kunnen worden en een heel groot
baar. Per ml ECS-cellen kunnen gemiddeld 20 tot
bladoppervlakte ontwikkelen. Aangezien na ge-
70 verschillende GGO-bananenplanten gegene-
netische transformatie heel veel planten moeten
reerd worden, afhankelijk van soort en de kwali-
geanalyseerd worden, kan deze analyse ook niet
teit van de ECS-cellen80, 109, 112-114.
op alle volgroeide planten uitgevoerd worden16.
Het biotechnologisch onderzoek op bananen
De tot nu toe ontwikkelde GGO-bananen ver-
ontstond in België, maar situeert zich momenteel
tonen resistentie tegen schimmels, virussen,
voornamelijk in Australië, België, Brazilië, China, In-
nematoden of bacteriën (zie verder). Deze resis-
dië, Kenia, Maleisië, Nigerië, Zuid-Afrika, Oeganda,
tenties worden steeds eerst aangetoond in vitro
Het Verenigd Koninkrijk en de Verenigde Staten4.
RNA-INTERFERENTIE (RNAI) ALS MIDDEL OM GENEN UIT TE SCHAKELEN101
De toenemende kennis over het erfelijk materiaal van organismen ligt aan de basis van nieuwe methoden om gewenste eigenschappen aan planten te geven. Zo kan RNA-interferentie (RNAi) gebruikt worden om zeer specifiek genen
uit te schakelen. Bij dit proces zijn twee types van erfelijk materiaal van zeer groot belang, DNA en RNA. Als virussen buiten beschouwing gelaten worden, kan DNA als universele drager van erfelijke informatie beschouwd worden.
Bij hogere organismen (plant en dier) bestaat DNA uit twee strengen en iedere cel van een welbepaald organisme bevat
hetzelfde DNA, doch iedere cel heeft niet dezelfde informatie nodig. Zo moet een wortelcel andere eiwitten aanmaken
dan een bladcel. Om van welbepaalde genetische informatie naar eiwitproductie te komen, zijn twee stappen nodig.
Eerst wordt het stukje DNA (gen) dat nodig is voor de productie van een eiwit overgeschreven (transcriptie) naar RNA.
Dit RNA verplaatst zich van de kern naar het cytoplasma van de cel waar het vervolgens vertaald wordt (translatie)
naar het overeenkomstige eiwit. Met RNAi wil men de transcriptie en/of translatie van bepaalde stukjes erfelijk materiaal uitschakelen. Indien het RNA afgebroken wordt vooraleer het kan vertaald worden, zal de boodschap vervat in
het DNA nooit tot de eiwitproductie leiden. Dit fenomeen noemt men “gene silencing”, het letterlijk stilleggen van de
genfunctie door het uitschakelen van het overeenkomstige RNA. Het verdedigingsmechanisme van de plant is gebaseerd op de aanwezigheid van dubbelstrengig RNA. In de plant is het boodschapper RNA (mRNA) opgebouwd uit één
streng. Wanneer viraal RNA in de plantencel vermeerderd wordt, zijn er dubbelstrengige RNA-moleculen aanwezig.
Dit wordt door de plantencel als niet-eigen beschouwd en ter verdediging breekt ze de dubbelstrengige RNA’s af. Dit
proces activeert een mechanisme dat ook alle gelijkaardige RNA-moleculen afbreekt, ook al komen ze voor als enkelstrengig. Deze afgebroken kleine RNA-stukjes komen vervolgens in contact met het mRNA van het doelwitgen, herkennen het, en zetten zich op dit mRNA vast. Dit is een signaal voor de afbraak van het geheel, met als gevolg dat het eiwit
waarvoor het doelwitgen codeert niet meer aangemaakt wordt. RNAi blijkt ook heel efficiënt in banaan te werken115.
Banaan, het groene goud van het Zuiden
Welke GGO-bananen bieden
toekomstperspectief?
nismen te introduceren in het bananengenoom.
Het betreft hier vooral genen die eiwitten of RNA
aanmaken die negatief inwerken op het metabolisme van de schimmel27, 117-122. Ze zorgen ofwel
Fusarium
voor de aanmaak van poriën in de schimmel-
oxysporum f. sp. cubense: De schimmel Fusa-
membraan waardoor de membraan afbreekt,
rium oxysporum vormt een ernstige bedreiging
of leggen belangrijke enzymen in de schimmel
voor de wereldwijde handel en productie van
stil. In beide situaties zal de schimmel sterven.
vele bananensoorten. Daarom proberen ver-
Wanneer de schimmel de bananenplant infec-
schillende wetenschappers via genetische modi-
teert, neemt hij gedurende korte tijd voedings-
ficatie bananenplanten te ontwikkelen die resis-
stoffen op uit de plantencel, maar als hij ook de
tentie vertonen tegen deze schimmel. Een eerste
toxische eiwitten opneemt, valt het metabolisme
manier om GGO-bananen te ontwikkelen die re-
stil, sterft de schimmel en stopt de infectie. Daar-
sistent zijn tegen Foc-TR1 en/of Foc-TR4 bestaat
om werden GGO-bananen ontwikkeld die kleine
erin om resistentiegenen vanuit “wilde” bananen
RNAi-moleculen (zie kaderstuk) aanmaken tegen
over te brengen naar de gewenste bananenva-
twee verschillende vitale genen van de schimmel,
riëteit. Een uitgebreide studie van acht verschil-
velvet en trancriptiefactor 127. Daarnaast hoopt
lende wilde bananenplanten toonde aan dat er
men ook dat genen die werkzaam zijn tegen de
. M. balbisiana
schimmel Fusarium oxysporum ook resistentie
vertoont de meeste symptomen, terwijl M. bas-
verlenen tegen andere schimmels, zoals Mycosp-
joo en M. itinerans volledig resistent zijn voor de
haerella fijiensis die Black Sigatoka veroorzaakt119.
schimmel. Enkel de genomen van M. balbisiana
De ultieme oplossing kan zelfs zijn om meerdere
en M. acuminata zijn echter ontrafeld (zie Hoofd-
genen binnen te brengen in één bananensoort
stuk 1). Omdat ook M. acuminata enige resisten-
die dan resistentie geven tegen verschillende
tie vertoont tegen Foc-TR4, betekent dit dus dat
ziekteverwekkers, want die zijn toch dikwijls al-
deze voorouder minstens één gen bezit dat ver-
lemaal samen aanwezig op het land123, 124. Tot nu
antwoordelijk is voor deze resistentie. Onderzoe-
toe werden de meeste resistentie-experimenten
kers zijn erin geslaagd dit gen of deze genen te
tegen Fusarium enkel in vitro en in de planten-
identificeren en met behulp van genetische trans-
kamer uitgetest29. Er zijn nog geen resultaten
formatie over te brengen naar het genoom van
beschikbaar van de Race1 veldproeven117-119 en
de ‘Cavendish’-banaan17. Na een eerste analyse
veldproeven voor Race 4-tolerantie zijn nog niet
in
uitgevoerd120-122.
GGO-bananen
resistent
tegen
een resistentievariatie bestaat
de
plantenkamer,
116
werden
een
aantal
GGO-planten opgegroeid op gronden besmet
met Fusarium oxysporum in Australië gedurende
een 18-maanden lange veldproef17. De resulta-
GGO-bananen resistent tegen Black Sigatoka:
ten van deze veldproef werden in de literatuur
Ook tegen de zeer destructieve bladvlekken-
nog niet gepubliceerd.
ziekte tracht men resistente bananenplanten te
ontwikkelen via gentechnologie. In 2013 bracht
Een tweede manier om resistentie tegen Fusari-
men genen uit rijst, rcc2 en rcg3, die instaan voor
um te bekomen, is om genen van andere orga-
de aanmaak van chitinase125, tot expressie in de
43
‘Gros Michel’-banaan16. Chitinase is een enyz-
ten komen van nature immers voor in een aantal
verder in een veldproef geanalyseerd worden op
brengststijging zullen nu ook getest worden voor
me dat chitine, een belangrijk bestanddeel van
andere planten, zoals rijst, tomaten en pepers,
hun resistentie tegen Xanthomonas campestris.
resistentie tegen andere nematoden, in het bij-
de celwand van schimmels, afbreekt. Wanneer
die rauw gegeten worden. GGO-planten met het
Ook andere belangrijke eigenschappen zoals fr-
zonder Meloidogyne spp. en Pratylenchus spp.20
de GGO-’Gros Michel’ het rijstchitinase produ-
Hrap- of het Pflp-gen werden gegenereerd voor
uitkwaliteit, zullen bestudeerd worden67.
ceert, wordt dit door de schimmel opgenomen
twee bananenvariëteiten, ‘Sukali ndiizi’ (AAB) en
bij infectie, en de celwand van de schimmel af-
‘Nakinyika’ (AAA). Veertig GGO-Hrap- en 25 GGO-
gebroken. Hierdoor wordt de schimmelinfectie
Pflp-planten vertoonden resistentie tegen de
tenietgedaan en is deze GGO-’Gros Michel’ resis-
bacterie in de plantenkamer en werden daarom
tent voor Black Sigatoka16. Men ontwikkelde 17
tijdens een veldproef in Oeganda getest126. Op
GGO-’Gros Michel’ bananenplanten, 2 rcc2 lijnen
het veld verschilden de GGO-bananenplanten
en 15 rcg 3 lijnen, en testte deze in vitro voor hun
morfologisch niet van hun niet-GGO-soortgeno-
resistentie tegen de M. fijensis schimmel. Negen
ten en hun groei en fruitontwikkeling waren nor-
van deze GGO-bananen vertoonden 39 dagen
maal. Vóór de bloei infecteerden de onderzoe-
na infectie nog geen ziektepatronen, en 108 da-
kers alle planten met Xanthomonas. Terwijl alle
gen na infectie (eindpunt van de studie) waren
niet-GGO-bananenplanten symptomen vertoon-
de necrose gebieden beperkt tot 16% in vergelij-
den en uiteindelijk verwelkten, bleken 20% van
king met de niet-getransformeerde plant16.
de Hrap-GGO-planten en 16% van de Pflp-GGOplanten geen ziektensymptomen te vertonen in
GGO-bananen resistent tegen BXW: Gezien de
vlugge verspreiding van BXW doorheen Afrika en
het gebrek aan resistentiegenen in banaan, is genetische transformatie de enige manier om weerstand te bieden tegen deze bacterie. Om BXW te
bestrijden werd het “Banana Bacterial Wilt” project opgericht met de bedoeling BXW-resistente
bananen te ontwikkelen voor de plaatselijke boeren. In dit project zijn zowel de "African Agricultural Technology Foundation" (AATF), IITA als NARO
betrokken. In een eerste studie werden twee
resistentiegenen gebruikt afkomstig van zoete
peper, nl. het Hrap-gen dat een eiwit dat planten beschermt tegen bacteriën aanmaakt en het
Pflp-gen, een ferrodoxine-achtig eiwit126, 127. Beide
genen hadden al hun werking tegen andere bacteriën, zoals Erwinia carotovora ssp. carotovora en
Pseudomonas syringae aangetoond in transgene
Arabidopsis thaliana en tabaksplanten128, 129. De
gecodeerde eiwitten staan ook niet op de lijst van
allergieverwekkers en kunnen dus voor menselijke consumptie toegelaten worden. Deze eiwit-
Banaan, het groene goud van het Zuiden
het veld126. Heel belangrijk was ook dat een resistente moederplant in staat was om de resistentie
door te geven aan de dochterplanten126. In de
toekomst wil men nu bananenplanten ontwikkelen die beide resistentiegenen, Hrap en Pflp, bezitten, waardoor het voor de bacterie nog moeilijker wordt om de bananenplant te infecteren. In
een tweede studie introduceerden de onderzoekers het XA21-gen van rijst in de bananenvariëteit
‘Gonja manjaya’ (AAB)67. Dit XA21-gen maakt rijst
resistent tegen infectie met Xanthomonas oryzae
en blijkt ook werkzaam te zijn tegen bacteriële
infecties in GGO-citrus en GGO-tomaat130, 131. In
vitro vertoonden 12/25 GGO-bananen resistentie
tegen Xanthomonas campestris cv. musacearum.
Deze resistente planten werden vervolgens overgebracht naar de plantenkamer waar hun resistentie op 90-dagen-oude plantjes nogmaals getest werden. De resistentie van de GGO-planten
varieerde, maar 58% van de planten waren volledig resistent. De XA21-expressie in banaan had
geen effect op de meetbare eigenschappen van
de plant. Deze resistente GGO-planten zullen nu
GGO-bananen tegen nematoden: Een mogelijke
GGO-bananen tegen snuitkevers: Er bestaat
aanpak om resistente bananenplanten tegen
heel wat controversie over de impact van snuit-
nematoden te ontwikkelen is het verterings-
kevers op de bananenteelt, die door sommigen
proces van de wormpjes te blokkeren tijdens
als een “belangrijke” of “ernstige” bedreiging be-
infectie van de plant. Belangrijke verteringsen-
schouwd wordt, terwijl anderen het eerder als
zymen bij nematoden zijn cysteine proteïna-
“lokaal”, “beperkt” of “niet belangrijk” aanzien61.
ses. In planten bestaan er proteïnase-inhibito-
Vandaar dat onderzoek naar resistenties eerder
ren, cystatines, die deze cysteine proteïnases
beperkt blijft. In de literatuur over natuurlijke re-
blokkeren, waardoor de nematode sterft. Op
sistentie tegen de snuitkever bij bananen zijn de
, Ara-
resultaten dikwijls tegenstrijdig61. Op een paar uit-
, rijst135, en aardappelen136 gege-
zonderingen na blijken de bakbananen (AAB) en
nereerd. Bij een tweede aanpak wordt op de
hooglandbananen (AAA-EA) meer gevoelig voor
nematodeninvasie in de bananenplant gewerkt
de kevers dan de andere genoomgroepen63. Van
door twee synthetische peptiden (een molecule
de twee voorouders blijkt de Musa acuminata (AA)
met een beperkt aantal aminozuren) tot expre-
gevoeliger te zijn dan de Musa balbisiana (BB)139.
sie te brengen in de wortels van de plant137, 138.
Tenslotte blijken diploïden resistenter te zijn dan
Hierbij wordt zowel het binnendringen van de
polyploïden140. Momenteel trachten vooral NARO,
nematode via de wortel, als de ontwikkeling van
IITA en “African Centre for Banana and Plantain”
de nematodenpopulatie in de wortel tegenge-
(CARBAP) in Cameroon snuitkever-resistente ba-
gaan. In GGO-bakbananen van de variëteit ‘Gon-
nanen te bekomen via veredeling63. Daarvoor
ja manjaya’ werden het cystatine van maïs, het
worden vooral de diploïde AA ouderplanten 'Cal-
synthetisch peptide of beide genen tot expressie
cutta4' en 'Pisang Lilin’ gebruikt140. Daarnaast is er
gebracht20, 114. Deze planten bleken in de serre re-
ook nood aan een eenvoudige en vlugge scree-
sistentie te vertonen tegen de nematode Radop-
ningsmethode om resistentie tegen de snuitke-
holus similis, tegen Helicotylenchus multicinctus en
vers in het laboratorium, groeikamers en op het
. Vervolgens
veld te testen. Tenslotte wordt de hulp van de
werd de resistentie in meer detail onderzocht
biotechnologie ingeroepen om GGO-bananen te
bij 12 GGO-planten tijdens een veldproef in Oe-
ontwikkelen die resistent zijn tegen de snuitke-
ganda20. Een duidelijke daling in de hoeveelheid
ver63, waarbij vooral op het spijsverteringsstelsel
nematoden werd bij alle GGO-planten waargeno-
van de kever wordt ingewerkt, meer bepaald door
men, waarvan acht GGO-planten significant resis-
het tot expressie brengen van een combinatie
tent waren tijdens de vegetatieve ontwikkeling,
van serine en cysteïne proteïnasen63. Dit onder-
10 op het bloeitijdstip en alle 12 op het oogsttijd-
zoek is de focus van een samenwerkingsverband
stip. Daarnaast steeg duidelijk de fruitopbrengst,
tussen NARO, IITA en “Forestry and Agricultural
die zelfs opliep tot 186% in vergelijking met de
Biotechnology Institute” (FABI) van de Universiteit
controleplant. De planten met de hoogste op-
van Pretoria, Zuid Afrika.
deze manier werden resistente tomaten
132
bidopsis133,
134
zelfs tegen een mengsel van beide
114
45
Ook bananen zijn een belangrijke bron van pVAC,
Bananen met meer pVAC kunnen op twee ma-
maar de hoeveelheden variëren heel sterk van
nieren bekomen worden: (1) via klassieke verede-
soort tot soort. Een uitvoerige analyse van 171
ling, en (2) via genetische modificatie. HarvestPlus
bananensoorten toonde aan de pVAC-hoeveel-
(http://www.ifpri.org/program/harvestplus) finan-
heden kunnen fluctueren van 0 tot 3500 µg/100 g
ciert het onderzoek dat via veredeling bananen
versgewicht , wat impliceert dat het verorberen
met hoge pVAC-hoeveelheden voor de bevolking
van één banaan van bepaalde soorten per dag
in Sub-Sahara-Afrika wenst te bekomen. Hierbij
reeds voldoende is om aan 50% van de dagelijk-
worden vooral bananen met elkaar gekruist die
se behoefte aan pVAC te voldoen. De bananen-
hoge hoeveelheden pVAC produceren, een hoge
soorten die in Oost-Afrika gegeten worden, zoals
opbrengst vertonen en ziekte-resistent zijn. Door-
de hoogland- en bakbananen, bevatten echter
dat de meeste (eetbare) bananen echter trïploid
pVAC hoeveelheden die variëren van 3,89 tot
en steriel zijn, is deze veredeling niet eenvoudig.
142
18,75 µg/g versgewicht143. Er is een correlatie tussen de kleur van het vruchtvlees en de hoeveel-
Sinds 2007 steunt de Bill en Melinda Gates Stich-
heid pVAC: bananen met een wit, crèmekleurig
ting een project waarbij onderzoekers uit Australië
vruchtvlees, zoals de ‘Cavendish’-banaan produ-
(Queensland University of Technology) en Oegan-
ceren zeer weinig pVAC, terwijl bij bananen met
da (NARO) GGO-bananen met verhoogde pVAC
donker oranje vruchtvlees, zoals de ‘Karat’ en de
en ijzergehaltes trachten te ontwikkelen. Hierbij
‘Asupina’-banaan, de pVAC hoeveelheden zeer
wordt het PSY2a gen van de ‘Asupina’-banaan, een
(www.promusa.org). Verder blijken
banaan met hoge pVAC-hoeveelheden, binnen-
de AA diploïden en de AAB en ABB triploïden ho-
gebracht in andere bananensoorten. Het PSY2a-
gere hoeveelheden pVAC te bevatten dan de AAA
gen codeert voor fytoeensynthase, een eiwit dat
bananen, maar toch kan er geen duidelijke cor-
belangrijk is bij de aanmaak van β-caroteen of
relatie aangetoond worden tussen genotype en
provitamine A. Dit gen komt in alle bananensoor-
hoeveelheid pVAC21, 145, 146. Wel lijken de pVAC-ni-
ten voor, maar het aangemaakte eiwit heeft in de
veaus in bakbananen hoger te zijn dan in kook- en
verschillende soorten een verschillende enzymati-
dessertbananen. Tenslotte varieert, afhankelijk
sche activiteit, waardoor sommige bananen, zoals
van de bananensoort, ook nog het pVAC-gehalte
de ‘Asupina’-banaan, meer pVAC bevatten dan an-
tussen onrijp en rijp fruit146.
dere bananen (zoals de ‘Cavendish’-banaan)144. De
hoog zijn
5
Banaantjes met extra vitamine A
In ontwikkelingslanden, en vooral in Sub-Sahara-Afrika en Zuidoost-India, kampen zeer
veel mensen met een vitamine A-tekort. Dit vitamine A-tekort treft vooral kinderen
onder de 5 jaar en zwangere vrouwen en kan tot blindheid, verminderde groei en
beenderenontwikkeling en zelfs tot de dood leiden. Meer dan 80% van de vitamine
A-opname door de bevolking in ontwikkelingslanden gebeurt via planten, onder de vorm
van provitamine A carotenoïden (pVAC) die in het menselijk lichaam vervolgens in retinol
en tenslotte in vitamine A omgezet worden 141.
(Opgroeien van bananenscheuten in Costa Rica (Bron: R. Swennen))
Banaan, het groene goud van het Zuiden
1, 144
werking van het additionele PSY2a gen wordt eerst
Ook de verwerking van bananen in voeding blijkt
uitgetest in de ‘Cavendish’ soorten ‘Williams’ en
een negatief effect te hebben op de hoeveelheid
‘Dwarf Cavendish’ en in de ‘Pome’ soort ‘Lady Fin-
pVAC. Verschillende processen, zoals blootstel-
ger’, maar het uiteindelijke doel is om de techno-
ling aan licht en lucht, beschadiging tijdens op-
logie over te brengen naar de hooglandbananen
slag en transport, invriezen, koken en frituren
en de ziekte-resistente hybride ‘Kabana 6H’, die
zorgen voor een daling in pVAC-hoeveelheden
146
na kruisingen bekomen werd147. In Australië (sinds
(www.Promusa.org), met als gevolg dat er meer
2008) en Oeganda (sinds 2010) werden reeds
vruchten per dag moeten geconsumeerd wor-
veldproeven met de GGO-’Cavendish’ bananen
den om aan de dagelijkse behoefte van vitamine
uitgevoerd en konden een aantal GGO-planten
A te voldoen.
met verhoogde pVAC geïdentificeerd worden147.
47
6 Besluit
Bananen zijn levensnoodzakelijk voor miljoenen mensen in het Zuiden. Niet alleen worden de bananen
verkocht op de lokale markten, waardoor ze een bron van inkomen zijn, maar zijn ze ook één van de
belangrijkste voedselbronnen. Bananen zijn ook de eerste gedomesticeerde gewassen ter wereld
maar vreemd genoeg gebruiken zowel de grote als kleine kwekers nauwelijks veredelde variëteiten.
De bananenteelt wordt echter door zeer veel ziektes en plagen bedreigd, die binnen de perken
kunnen gehouden worden door een correcte landbouwstrategie of door het ontwikkelen van nieuwe
variëteiten via veredeling. Voor sommige bedreigingen kunnen enkel GGO-bananen een duurzame
en stabiele oplossing bieden. In het geval van de bananen is, ondanks soms sterke tegenkantingen
van bepaalde NGO’s, het gebruik van gentechnologie zeker verantwoord. Populaire bananensoorten
kunnen immers dikwijls op geen enkele andere manier verbeterd worden, aangezien resistentiegenen
vaak niet aanwezig zijn bij bananen. Hierdoor moet naar resistentiegenen van andere plantensoorten
of zelfs andere organismen gezocht worden. Daarnaast voldoen bananen, bekomen na veredeling,
vaak niet aan de eisen van de consument qua kleur, smaak, of andere voorname eigenschappen. Een
belangrijke argumentatie van tegenstanders van de GGO-technologie, namelijk dat de transgenen zich
ongecontroleerd kunnen verspreiden in de natuur, is bij de banaan niet van toepassing. Aangezien
de meeste bananensoorten triploïd, zaadloos en steriel zijn, zullen de nieuwe genen, eenmaal ze
geïncorporeerd zijn in het bananengenoom, niet meer via het pollen verspreid kunnen worden en dus
niet ongecontroleerd in de natuur terechtkomen. Wanneer geen GGO-bananen aanvaard worden,
dreigen zowel de bananenindustrie als de lokale voedselvoorziening in Afrika volledig stil te vallen. Een
analoge situatie deed zich voor in de jaren 1990 bij de papaja-industrie op Hawaï. Infecties van papaja
met het “papaja ringspot”-virus bedreigde de volledige fruitproductie. De publieke sector ontwikkelde
een virus-resistente GGO-papaja. De zaden werden in 1998 op de markt gebracht en in 2000 stond
het papaja-productieniveau in Hawaï terug op het peil van voor de uitbraak van het virus101. Alle GGObananen zijn momenteel triploïd. In de toekomst is het ook wenselijk dat er diploïde GGO-bananen,
dus met twee sets chromosomen, ontwikkeld worden. Diploïden kunnen immers veel gemakkelijker
gekruist worden, waardoor de GGO-technologie de verdere ontwikkeling van nieuwe variëteiten via
veredeling zal kunnen ondersteunen.
Banaan, het groene goud van het Zuiden
49
7
Referenties
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
Pereira, A., Maraschin, M. (2015). Banana (Musa spp) from peel to pulp: Ethnopharmacology, source of bioactive compounds and
its relevance for human health. J Ethnopharmacol 160: 149–163.
Janssens, S.B., Vandelook, F., De Langhe, E., Verstraete, B., Smets, E., Vandenhouwe, I., Swennen, R. (2016). Evolutionary dynamics
and biogeography of Musaceae reveal a correlation between the diversification of the banana family and the geological and
climatic history of Southeast Asia. New Phytol. DOI: 10.1111/nph.13856.
Perrier, X., De Langhe, E., Donohue, M., Lentfer, C., Vrydaghs, L., Bakry, F., Carreel, F., Hippolyte, I., Horry, J.P., Jenny, C., Lebot, V.,
Risterucci, A.M., Tomekpe, K., Doutrelepont, H., Ball, T., Manwaring, J., de Maret, P., Denham, T. (2011).
Multidisciplinary perspectives on banana (Musa spp.) domestication. P Natl Acad Sci USA 108: 11311–11318.
Ortiz, R., Swennen, R. (2014). From crossbreeding to biotechnology-facilitated improvement of banana and plantain.
Biotechnol Adv 32: 158–169.
Valmayor, R.V. (2000). Cooking bananas - Classification, production and utilization in Southeast Asia. Infomusa 9: 28–30.
Taylor, J.S., Erkek, E. (2004). Latex allergy: diagnosis and management. Dermatologic Therapy 17: 289–301.
Passo Tsamo, C.V., Herent, M.F., Tomekpe, K., Happi Emaga, T., Quetin-Leclercq, J., Rogez, H., Larondelle, Y., Andre, C. (2015).
Phenolic profiling in the pulp and peel of nine plantain cultivars (Musa sp.). Food Chem 167: 197–204.
Davey, M.W., Gudimella, R., Harikrishna, J.A., Sin, L.W., Khalid, N., Keulemans, J. (2013). “A draft Musa balbisiana genome sequence
for molecular genetics in polyploid, inter- and intra-specific Musa hybrids”. Bmc Genomics 14: 683.
McKey, D., Elias, M., Pujol, B., Duputie, A. (2010). The evolutionary ecology of clonally propagated domesticated plants.
New Phytol 186: 318–332.
Simmonds, N.W., Shepherd, K. (1955). The taxonomy and origins of the cultivated bananas.
Journal of the Linnean Society of London, Botany 55: 302–312.
Heslop-Harrison, J.S., Schwarzacher, T. (2007). Domestication, genomics and the future for banana.
Ann Bot-London 100: 1073–1084.
El-Khishin, D.A., Belatus, E.L., El-Hamid, A.A., Radwan, K.H. (2009). Molecular characterization of banana cultivars (Musa spp.) from
Egypt using AFLP. Research Journal of Agriculture and Biological Sciences 5: 272–279.
D’Hont, A., Denoeud, F., Aury, J.M., Baurens, F.C., Carreel, F., Garsmeur, O., Noel, B., Bocs, S., Droc, G., Rouard, M., Da Silva, C.,
Jabbari, K., Cardi, C., Poulain, J., Souquet, M., Labadie, K., Jourda, C., Lengellé, J., Rodier-Goud, M., Alberti, A., Bernard, M.,
Correa, M., Ayyampalayam, S., Mckain, M.R., Leebens-Mack, J., Burgess, D., Freeling, M., Mbéguié-A-Mbéguié, D., Chabannes, M.,
Wicker, T., Panaud, O., Barbosa, J., Hribova, E., Heslop-Harrison, P., Habas, R., Rivallan, R., Francois, P., Poiron, C., Kilian, A., Burthia,
D., Jenny, C., Bakry, F., Brown, S., Guignon, V., Kema, G., Dita, M., Waalwijk, C., Joseph, S., Dievart, A., Jaillon, O., Leclercq, J., Argout,
X., Lyons, E., Almeida, A., Jeridi, M., Dolezel, J., Roux, N., Risterucci, A.M., Weissenbach, J., Ruiz, M., Glaszmann, J.C., Quétier,
F., Yahiaoui, N., Wincker, P. (2012). The banana (Musa acuminata) genome and the evolution of monocotyledonous plants.
Nature 488: 213–218.
FAO (2015). FAOSTAT. Available at http:/faostat3.fao.org/home/index.html#DOWNLOAD.
Ploetz, R.C., Kema, G.H., Ma, L.J. (2015). Impact of diseases on export and smallholder production of banana.
Annu Rev Phytopathol 53: 269–288.
Kovács, G., Sági, L., Jacon, G., Arinaitwe, G., Busogoro, J.P., Thiry, E., Strosse, H., Swennen, R., Remy, S. (2013). Expression of a rice
chitinase gene in transgenic banana (‘Gros Michel’, AAA genome group) confers resistance to Black Leaf Streak disease.
Transgenic Res 22: 117–130.
Butler, D. (2013). Fungus threatens top banana. Nature 504: 195–196.
Ordóñez, N., García-Bastidas, F., Laghari, H.B., Akkary, M.Y., Harfouche, E.N., al Awar, B.N., Kema, G.H.J. (2015). First report of
Fusarium oxysporum f. sp. cubense tropical race 4 causing panama disease in cavendish bananas in Pakistan and Lebanon.
Plant Dis 100: 209.
Atkinson, H.J., Roderick, H., Tripathi, L. (2015). Africa needs streamlined regulation to support the deployment of GM crops.
Trends Biotechnol 33: 433–435.
Tripathi, L., Babirye, A., Roderick, H., Tripathi, J.N., Changa, C., Urwin, P.E., Tushemereirwe, W.K., Coyne, D., Atkinson, H.J. (2015).
Field resistance of transgenic plantain to nematodes has potential for future African food security. Sci Rep-Uk 5.
Englberger, L., Darnton-Hill, I., Coyne, T., Fitzgerald, M.H., Marks, G.C. (2003). Carotenoid-rich bananas: a potential food source for
alleviating vitamin A deficiency. Food and nutrition bulletin 24: 303–318.
Tripathi, L., Mwangi, M., Abele, S., Aritua, V., Tushemereirwe, W.K., Bandyopadhyay, R. (2009). Xanthomonas wilt: A threat to
banana production in East and Central Africa. Plant Dis 93: 440–451.
Holmes, B. (2013). Go, bananas. New Scientist 218: 38–41.
Norgrove, L., Hauser, S. (2014). Improving plantain (Musa spp. AAB) yields on smallholder farms in West and Central Africa.
Food Secur 6: 501–514.
Michielse, C.B., Rep, M. (2009). Pathogen profile update: Fusarium oxysporum. Mol Plant Pathol 10: 311–324.
O’Donnell, K., Gueidan, C., Sink, S., Johnston, P.R., Crous, P.W., Glenn, A., Riley, R., Zitomer, N.C., Colyer, P., Waalwijk,
C., van der Lee, T., Moretti, A., Kang, S., Kim, H.S., Geiser, D.M., Juba, J.H., Baayen, R.P., Cromey, M.G., Bithell, S., Sutton, D.A.,
Skovgaard, K., Ploetz, R., Kistler, H.C., Elliott, M., Davis, M., Sarver, B.A.J. (2009). A two-locus DNA sequence database for typing
plant and human pathogens within the Fusarium oxysporum species complex. Fungal Genet Biol 46: 936–948.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
Ghag, S.B., Shekhawat, U.K.S., Ganapathi, T.R. (2014). Host-induced post-transcriptional hairpin RNA-mediated gene silencing of
vital fungal genes confers efficient resistance against Fusarium wilt in banana. Plant Biotechnol J 12: 541–553.
Buddenhagen, I. (2009). Understanding strain diversity in Fusarium oxysporum f. sp cubense and history of introduction of
‘Tropical Race 4’ to better manage banana production. Acta Hortic 828: 193–204.
Ploetz, R.C. (2015). Management of Fusarium wilt of banana: A review with special reference to tropical race 4. Crop Prot 73: 7–15.
Stover, R.H. (1986). Disease management strategies and the survival of the banana industry.
Annual Review of Phytopathology 24: 83–91.
Buddenhagen, I.W. (1990). Banana breeding and Fusarium wilt. Fusarium Wilt of Banana: 107–113.
García-Bastidas, F., Ordóñez, N., Konkol, J., Al-Qasim, M., Naser, Z., Abdelwali, M., Salem, N., Waalwijk, C., Ploetz, R.C., Kema, G.H.J.
(2014). First Report of Fusarium oxysporum f. sp cubense Tropical Race 4 associated with panama disease of banana outside
Southeast Asia. Plant Dis 98: 694.
Ploetz, R.C. (2009). Assessing threats posed by destructive banana pathogens. Acta Hortic 828: 245–251.
Ploetz, R.C., Churchill, A.C.L. (2011). Fusarium wilt: the banana disease that refuses to go away. International Ishs-Promusa
Symposium on Global Perspectives on Asian Challenges 897: 519–526.
Meldrum, R.A., Daly, A.M., Tran-Nguyen, L.T.T., Aitken, E.A.B. (2013). Are banana weevil borers a vector in spreading Fusarium
oxysporum f. sp cubense tropical race 4 in banana plantations? Australas Plant Path 42: 543–549.
Ploetz, R.C. (2001). Black Sigatoka of banana: The most important disease of a most important fruit. The Plant Health Instructor.
de Bellaire, L.D., Foure, E., Abadie, C., Carlier, J. (2010). Black Leaf Streak Disease is challenging the banana industry.
Fruits 65: 327–342.
Saraiva, L.D., Castelan, F.P., Shitakubo, R., Hassimotto, N.M.A., Purgatto, E., Chillet, M., Cordenunsi, B.R. (2013). Black Leaf Streak
Disease affects starch metabolism in banana fruit. J Agr Food Chem 61: 5582–5589.
Bennett, R.S., Arneson, P.A. (2003). Black Sigatoka of bananas and plantains. The Plant Health Instructor.
Kumar, P.L., Selvarajan, R., Iskra-Caruana, M.-L., Chabannes, M., Hanna, R. (2015). Biology, Etiology, and Control of Virus Diseases
of Banana and Plantain. in Advances in Virus Research, L. Gad, I. K. Nikolaos, Eds. (Academic Press, 2015),
vol. Volume 91, pp. 229–269.
Elayabalan, S., Subramaniam, S., Selvarajan, R. (2015). Banana bunchy top disease (BBTD) symptom expression in banana and
strategies for transgenic resistance: A review. Emir J Food Agr 27: 55–74.
Hu, J.S., Wang, M., Sether, D., Xie, W., Leonhardt, K.W. (1996). Use of polymerase chain reaction (PCR) to study transmission of
banana bunchy top virus by the banana aphid (Pentalonia nigronervosa). Ann Appl Biol 128: 55–64.
Nelson, S.C. (2004). Banana bunchy top: Detailed signs and symptoms.
Available at http://www.ctahr.hawaii.edu/bbtd/downloads/bbtv-details.pdf.
Magee, C.J.P. (1927). Investigations of the bunchy top disease of bananas.
Available at http://www.musalit.org/seeMore.php?id=12192
Fahmy, T. (1927). Plant disease in Egypt. Minerals and agriculture in Egypt. Bulletin 30.
Blomme, G., Ploetz, R., Jones, D., De Langhe, E., Price, N., Gold, C., Geering, A., Viljoen, A., Karamura, D., Pillay, M., Tinzaara,
W., Teycheney, P.Y., Lepoint, P., Karamura, E., Buddenhagen, I. (2013). A historical overview of the appearance and spread
of Musa pests and pathogens on the African continent: highlighting the importance of clean Musa planting materials and
quarantine measures. Ann Appl Biol 162: 4–26.
Kumar, V., Campbell, L.M., Rathore, K.S. (2011). Rapid recovery- and characterization of transformants following Agrobacteriummediated T-DNA transfer to sorghum. Plant Cell Tiss Org 104: 137–146.
Magnaye, L.V., Espino, R.R.C. (1990). Note: Banana bract mosaic, a new disease of banana I. Symptomatology.
Philippine Agriculturist 73: 55–59.
Quito-Avila, D.R., Ibarra, M.A., Alvarez, R.A., Ratti, M.F., Espinoza, L., Cevallos-Cevallos, J.M., Peralta, E.L. (2013).
First report of banana bract mosaic virus in ‘Cavendish’ banana in Ecuador. Plant Dis 97: 1003.
Tushemereirwe, W., Kangire, A., Ssekiwoko, F., Offord, L.C., Crozier, J., Boa, E., Rutherford, M., Smith, J.J. (2004).
First report of Xanthomonas campestris pv. musacearum on banana in Uganda. Plant Pathol 53: 802.
Yirgou, D., Bradbury, J.F. (1968). Bacterial wilt of Enset (Ensete Ventricosum) incited by Xanthomonas musacearum sp
n. Phytopathology 58: 111–112.
Eden-Green, S. (2004). Focus on bacterial wilt. How can the advance of banana Xanthomonas wilt be halted? Infomusa 13: 38–41.
Tripathi, L., Odipio, J., Tripathi, J.N., Tusiime, G. (2008). A rapid technique for screening banana cultivars for resistance to
Xanthomonas wilt. Eur J Plant Pathol 121: 9–19.
Mwebaze, J.M., Tusiime, G., Tushemereirwe, W.K., Kubiriba, J. (2006). The survival of Xanthomonas campestris pv. musacearum in
soil and plant debris. African Crop Science Journal 14: 121–127.
Álvarez, E., Pantoja, A., Ceballos, G., Gañán, L. (2015). Managing Moko disease of banana in Latin America and the Caribbean.
Available at http://ciat-library.ciat.cgiar.org/Articulos_Ciat/biblioteca/Managing_Moko_Disease_of_Banana.pdf.
Brooks, F.E. (2008). Burrowing nematode. The Plant Health Instructor.
Coyne, D.L., Omowumi, A., Rotifa, I., Afolami, S.O. (2013). Pathogenicity and damage potential of five species of plant-parasitic
nematodes on plantain (Musa spp., AAB genome) cv. Agbagba. Nematology 15: 589–599.
De Luca, F., Troccoli, A., Duncan, L.W., Subbotin, S.A., Waeyenberge, L., Coyne, D.L., Brentu, F.C., Inserra, R.N. (2012). Pratylenchus
speijeri n. sp (Nematoda: Pratylenchidae), a new root-lesion nematode pest of plantain in West Africa. Nematology 14: 987–1004.
Niere, B., Gold, C.S., Coyne, D., Dubois, T., Sikora, R. (2014). Performance of tissue-cultured versus sucker-derived East African
highland banana (Musa AAA-EA) under high and low input systems in Uganda. Field Crop Res 156: 313–321.
Price, N.S. (2006). The banana burrowing nematode, Radopholus similis (Cobb) Thorne, in the Lake Victoria region of East Africa:
its introduction, spread and impact. Nematology 8: 801–817.
51
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
Gold, C., Pena, J., Karamura, E. (2001). Biology and integrated pest management for the banana weevil Cosmopolites sordidus
(Germar) (Coleoptera: Curculionidae). Integrated Pest Management Reviews 6: 79–155.
Gold, C.S., Bagabe, M.I., Ssendege, R. (1999). Banana weevil, Cosmopolites sordidus (Germar) (Coleoptera : Curculionidae):
tests for suspected resistance to carbofuran and dieldrin in the Masaka District, Uganda. Afr Entomol 7: 189–196.
Kiggundu, A., Pillay, M., Viljoen, A., Gold, C., Tushemereirwe, W., Kunert, K. (2003). Enhancing banana weevil (Cosmopolites sordidus)
resistance by plant genetic modification: A perspective. African Journal of Biotechnology 2: 563–569.
McIntyre, B., Gold, C., Kashaija, I., Ssali, H., Night, G., Bwamiki, D. (2001). Effects of legume intercrops on soil-borne pests, biomass,
nutrients and soil water in banana. Biol Fertil Soils 34: 342–348.
Pocasangre, L.E., Ploetz, R.C., Molina, A.B., Vicente, L.P. (2011). Raising Awareness of the Threat of Fusarium Wilt Tropical Race 4 in
Latin America and the Caribbean. International Ishs-Promusa Symposium on Global Perspectives on Asian Challenges 897: 331–337.
Kagezi, G.H., Kangire, A., Tushemereirwe, W., Bagamba, F., Kikulwe, E., Muhangi, J., Gold, C.S., Ragama, P. (2006). Banana bacterial
wilt incidence in Uganda. African Crop Science Journal 14: 83–91.
Tripathi, J.N., Lorenzen, J., Bahar, O., Ronald, P., Tripathi, L. (2014). Transgenic expression of the rice Xa21 pattern-recognition
receptor in banana (Musa sp.) confers resistance to Xanthomonas campestris pv. musacearum. Plant Biotechnol J 12: 663–673.
Njuguna, J., Nguthi, S., Wambugu, F., Gitau, D., Karuoya, M. (2011). Enhancing Banana Crop Management by Use of Tissue-Culture
Derived Planting Material in Kenya. International Ishs-Promusa Symposium on Global Perspectives on Asian Challenges 897: 459–462.
Lule, M., Dubois, T., Coyne, D., Kisitu, D., Kamusiime, H., Bbemba, J. (2013). Trainer”s manual. A training course for banana farmers
interested in growing tissue culture bananas. Available at http://www.iita.org/c/document_library/get_file?uuid=961e89b0-182743f5-bc8c-3f3fdf04ec10&groupId=25357.
Lule, M., Dubois, T., Coyne, D., Kisitu, D., Kamusiime, H., Bbemba, J. (2013). Trainer’s manual. A training course on setting up and
running a banana tissue culture nursery.
Available at http://www.iita.org/c/document_library/get_file?uuid=bebf6706-60a3-4d79-96fc-4a2f7c0da084&groupId=25357.
Smith, M.K., Whiley, A.W., Searle, C., Langdon, P.W., Schaffer, B., Pegg, K.G. (1998). Micropropagated bananas are more susceptible
to fusarium wilt than plants grown from conventional material. Aust J Agr Res 49: 1133–1139.
Köberl, M., Dita, M., Martinuz, A., Staver, C., Berg, G. (2015). Agroforestry leads to shifts within the gammaproteobacterial
microbiome of banana plants cultivated in Central America. Front Microbiol 6.
Dowiya, N.B., Rweyemamu, C.L., Maerere, A.P. (2009). Banana (Musa spp. Colla) cropping systems, production constraints and
cultivar preferences in eastern Democratic Republic of Congo. Journal of Animal & Plant Sciences 4: 341–356.
Muliele, M.T., Bielders, C.L., van Asten, P.J.A. (2015). Short- and medium-term impact of manual tillage and no-tillage with mulching
on banana roots and yields in banana-bean intercropping systems in the East African Highlands. Field Crop Res 171: 1–10.
Wang, B., Li, R., Ruan, Y., Ou, Y., Zhao, Y., Shen, Q. (2015). Pineapple–banana rotation reduced the amount of Fusarium oxysporum
more than maize–banana rotation mainly through modulating fungal communities. Soil Biology and Biochemistry 86: 77–86.
Meredith, D.S. (1943). The antagonism of Actinomyces to Fusarium oxysporum f. sp. cubense. Phytopathology 33: 403.
Lakshmanan, P., Selvaraj, P., Mohan, S. (1987). Efficacy of different methods for the control of Panama disease.
Trop Pest Manage 33: 373–376.
Nel, B., Steinberg, C., Labuschagne, N., Viljoen, A. (2007). Evaluation of fungicides and sterilants for potential application in the
management of Fusarium wilt of banana. Crop Prot 26: 697–705.
Herbert, J.A., Marx, D. (1990). Short-term control of Panama disease of bananas in South Africa. Phytophylactica 22: 339–340.
Remy, S., Kovacs, G., Swennen, R., Panis, B. (2013). Genetically Modified Bananas: Past, Present and Future.
Ii Genetically Modified Organisms in Horticulture Symposium 974: 71-80.
Marín, D.H., Romero, R.A., Guzmán, M., Sutton, T.B. (2003). Black Sigatoka: An increasing threat to banana cultivation.
Plant Dis 87: 208–222.
Candido, V., D’Addabbo, T., Basile, M., Castronuovo, D., Miccolis, V. (2008). Greenhouse soil solarization: effects on weeds,
nematodes and yield of tomato and melon. Agron Sustain Dev 28: 221–230.
zum Felde, A., Mendoza, A., Cabrera, J.A., Kurtz, A., Schouten, A., Pocasangre, L., Sikora, R.A. (2009). The Burrowing Nematode of
Banana: Strategies for Controlling the Uncontrollable. Acta Hortic 828: 101–107.
López-Lima, D., Sánchez-Nava, P., Carrión, G., Núñez-Sánchez, A.E. (2013). 89% reduction of a potato cyst nematode population
using biological control and rotation. Agron Sustain Dev 33: 425–431.
Ploetz, R., Freeman, S., Konkol, J., Al-Abed, A., Naser, Z., Shalan, K., Barakat, R., Israeli, Y. (2015). Tropical race 4 of Panama disease
in the Middle East. Phytoparasitica 43: 283-293.
Jesus, F.N., Damasceno, J.C.A., Barbosa, D.H.S.G., Malheiro, R., Pereira, J.A., Soares, A.C.F. (2015). Control of the banana burrowing
nematode using sisal extract. Agron Sustain Dev 35: 783–791.
El-Ansary, M.S.M., Hamouda, R.A. (2014). Biocontrol of root-knot nematode infected banana plants by some marine algae.
Russian Journal of Marine Biology 40: 140-146.
Yang, X., Wang, X., Wang, K., Su, L., Li, H., Li, R., Shen, Q. (2015). The nematicidal effect of Camellia seed cake on root-knot
nematode Meloidogyne javanica of banana. Plos One 10: e0119700.
Lorenzen, J., Tenkouano, A., Bandyopadhyay, R., Vroh, B., Coyne, D., Tripathi, L. (2010). Overview of banana and plantain
(Musa spp.) improvement in Africa: past and future Acta Horticulturae 879: 595–604.
Heslop-Harrison, J.S. (2011). Genomics, banana breeding and superdomestication. 55-62.
Ortiz, R. (2013). Conventional Banana and Plantain Breeding. International Ishs-Promusa Symposium on Bananas and Plantains:
Towards Sustainable Global Production and Improved Use 986: 177–194.
De Steur, H., Blancquaert, D., Strobbe, S., Lambert, W., Gellynck, X., Van Der Straeten, D. (2015). Status and market potential of
transgenic biofortified crops. Nat Biotechnol 33: 25–29.
93.
94.
95.
96.
97.
98.
99.
100.
101.
102.
103.
104.
105.
106.
107.
108.
109.
110.
111.
112.
113.
114.
115.
116.
117.
118.
119.
120.
121.
122.
123.
Shimelis, H., Laing, M. (2012). Timelines in conventional crop improvement: pre-breeding and breeding procedures.
Australian Journal of Crop Science 6: 1542–1549.
Swennen, R., Vuylsteke, D. (1993). Breeding Black Sigatoka resistant plantains with a wild banana. Trop Agr 70: 74–77.
Tenkouano, A., Swennen, R. (2004). Progress in breeding and delivering improved plantain and banana to African farmers.
Chronica Horticulturae 44: 9–15.
Tushemereirwe, W., Batte, M., Nyine, M., Tumuhimbise, R., Barekye, A., Tendo, S., Talengera, D., Kubiriba, J., Lorenzen, J., Swennen, R.,
Uwimana, B. (2015). Performance of Narita banana hybrids in the preliminary yield trial for three cycles in Uganda.
Available on: http://www.iita.org/c/document_library/get_file?uuid=1500c39e-7c05-4219-b649-85cdb1bd73f6&groupId=25357.
Dochez, C., Tenkouano, A., Ortiz, R., Whyte, J., De Waele, D. (2009). Host plant resistance to Radopholus similis in a diploid banana
hybrid population. Nematology 11: 329–335.
Quénéhervé, P., Valette, C., Topart, P., du Montcel, H.T., Salmon, F. (2009). Nematode resistance in bananas: screening results on
some wild and cultivated accessions of Musa spp. Euphytica 165: 123–136.
Vaughan, D.A., Balázs, E., Heslop-Harrison, J.S. (2007). From crop domestication to super-domestication.
Ann Bot-London 100: 893–901.
VIB (2014). Een schimmelresistente aardappel voor België..
Available at http://www.vib.be/en/about-vib/plant-biotech-news/Documents/BackgroundReport_Potato_ENG.pdf
VIB (2014). Virus resistente Papaja in Hawaï.
Available at http://www.vib.be/en/about-vib/plant-biotech-news/Documents/Virus%20resistant%20papaya%20in%20Hawaii.pdf
Kyndt, T., Quispe, D., Zhai, H., Jarret, R., Ghislain, M., Liu, Q., Gheysen, G., Kreuze, J.F. (2015). The genome of cultivated sweet potato
contains Agrobacterium T-DNAs with expressed genes: An example of a naturally transgenic food crop.
P Natl Acad Sci USA 112: 5844–5849.
Sági, L., Panis, B., Remy, S., Schoofs, H., De Smet, K., Swennen, R., Cammue, B.P.A. (1995). Genetic transformation of banana and
plantain (Musa spp) via particle bombardment. Bio-Technol 13: 481–485.
Panis, B., Van Wauwe, A., Swennen, R. (1993). Plant regeneration through direct somatic embryogenesis from protoplasts of
banana (Musa spp). Plant Cell Rep 12: 403–407.
May, G.D., Afza, R., Mason, H.S., Wiecko, A., Novak, F.J., Arntzen, C.J. (1995). Generation of transgenic banana (Musa acuminata)
plants via Agrobacterium-mediated transformation. Bio-Technol 13: 486–492.
Sagi, L., Remy, S., Cammue, B.P., Maes, K., Raemaeckers, T., Panis, B., Schoofs, H., Swennen, R. (2000). Production of transgenic
banana and plantain. Acta Horticulturae 540: 203–206.
Becker, D.K., Dugdale, B., Smith, M.K., Harding, R.M., Dale, J.L. (2000). Genetic transformation of Cavendish banana (Musa spp.
AAA group) cv ‘Grand Nain’ via microprojectile bombardment. Plant Cell Rep 19: 229–234.
Ganapathi, T.R., Higgs, N.S., Balint-Kurti, P.J., Arntzen, C.J., May, G.D., Van Eck, J.M. (2001). Agrobacterium-mediated transformation of
embryogenic cell suspensions of the banana cultivar Rasthali (AAB). Plant Cell Rep 20: 157–162.
Tripathi, J.N., Oduor, R.O., Tripathi, L. (2015). A high-throughput regeneration and transformation platform for production of
genetically modified banana. Frontiers in Plant Science 6.
Strosse, H., Schoofs, H., Panis, B., Andre, E., Reyniers, K., Swennen, R. (2006). Development of embryogenic cell suspensions from
shoot meristematic tissue in bananas and plantains (Musa spp.). Plant Sci 170: 104–112.
Tripathi, J.N., Muwonge, A., Tripathi, L. (2012). Efficient regeneration and transformation of plantain cv. “Gonja manjaya”
(Musa spp. AAB) using embryogenic cell suspensions. In Vitro Cell Dev-Pl 48: 216–224.
Pérez-Hernández, J., Swennen, R., Sági, L. (2006). Number and accuracy of T-DNA insertions in transgenic banana (Musa spp.)
plants characterized by an improved anchored PCR technique. Transgenic Res 15: 139–150.
Chong-Pérez, B., Reyes, M., Rojas, L., Ocaña, B., Perez, B., Kosky, R.G., Angenon, G. (2012). Establishment of embryogenic cell
suspension cultures and Agrobacterium-mediated transformation in banana cv. ‘Dwarf Cavendish’ (Musa AAA): effect of spermidine
on transformation efficiency. Plant Cell Tiss Org 111: 79–90.
Roderick, H., Tripathi, L., Babirye, A., Wang, D., Tripathi, J., Urwin, P.E., Atkinson, H.J. (2012). Generation of transgenic plantain
(Musa spp.) with resistance to plant pathogenic nematodes. Mol Plant Pathol 13: 842–851.
Dang, T.V.T., Windelinckx, S., Henry, I.M., De Coninck, B., Cammue, B.P.A., Swennen, R., Remy, S. (2014). Assessment of RNAiinduced silencing in banana (Musa spp.). BMC Research Notes 7: 655.
Li, W.M., Dita, M., Wu, W., Hu, G.B., Xie, J.H., Ge, X.J. (2015). Resistance sources to Fusarium oxysporum f. sp cubense tropical race 4 in
banana wild relatives. Plant Pathol 64: 1061-1067.
Subramaniam, S., Maziah, M., Sariah, M., Puad, M.P., Xavier, R. (2006). Bioassay method for testing Fusarium wilt disease tolerance
in transgenic banana. Sci Hortic-Amsterdam 108: 378–389.
Paul, J., Becker, D.K., Dickman, M.B., Harding, R.M., Khanna, H.K., Dale, J.L. (2011). Apoptosis-related genes confer resistance to
fusarium wilt in transgenic ‘lady finger’ bananas. Plant Biotech J 9.
Ghag, S.B., Shekhawat, U.K.S., Ganapathi, T.R. (2012). Petunia floral defensins with unique prodomains as novel candidates for
development of Fusarium wilt resistance in transgenic banana plants. Plos One 7: e39557.
Hu, C.-H., Wei, Y.-R., Huang, Y.-H., Yi, G.-J. (2013). An efficient protocol for the production of chit42 transgenic Furenzhi banana
(Musa spp. AA group) resistant to Fusarium oxysporum. In Vitro Cell Dev-Pl 49: 584–-592.
Mahdavi, F., Sariah, M., Maziah, M. (2012). Expression of rice thaumatin-like protein gene in transgenic banana plants enhances
resistance to Fusarium wilt. Appl Biochem Biotech 166: 1008–1019.
Yip, M.-K., Lee, S.-W., Su, K.-C., Lin, Y.-H., Chen, T.-Y., Feng, T.-Y. (2011). An easy and efficient protocol in the production of pflp
transgenic banana against Fusarium wilt. Plant Biotechnol Rep 5: 245–254.
Halpin, C. (2005). Gene stacking in transgenic plants - the challenge for 21st century plant biotechnology. Plant Biotechnol J 3: 141–155.
53
124. Remy, S., François, I., Cammue, B.P.A., Swennen, R., Sági, L. (1998). Co-transformation as a potential tool to create multiple and
durable resistance in banana (Musa spp.). Acta Hort 461.
125. Nishizawa, Y., Nishio, Z., Nakazono, K., Soma, M., Nakajima, E., Ugaki, M., Hibi, T. (1999). Enhanced resistance to blast
(Magnaporthe grisea) in transgenic Japonica rice by constitutive expression of rice chitinase. Theor Appl Genet 99: 383–390.
126. Tripathi, L., Mwaka, H., Tripathi, J.N., Tushemereirwe, W.K. (2010). Expression of sweet pepper Hrap gene in banana enhances
resistance to Xanthomonas campestris pv. musacearum. Mol Plant Pathol 11: 721–731.
127. Namukwaya, B., Tripathi, L., Tripathi, J.N., Arinaitwe, G., Mukasa, S.B., Tushemereirwe, W.K. (2012). Transgenic banana expressing
Pflp gene confers enhanced resistance to Xanthomonas wilt disease. Transgenic Res 21: 855–865.
128. Ger, M.-J., Chen, C.-H., Hwang, S.-Y., Huang, H.-E., Podile, A.R., Dayakar, B.V., Feng, T.-Y. (2002). Constitutive expression of hrap
gene in transgenic tobacco plant enhances resistance against virulent bacterial pathogens by induction of a hypersensitive
response. Mol Plant Microbe In 15: 764–773.
129. Pandey, A.-K., Ger, M.-J., Huang, H.-E., Yip, M.-K., Zeng, J., Feng, T.-Y. (2005). Expression of the hypersensitive response-assisting
protein in Arabidopsis results in harpin-dependent hypersensitive cell death in response to Erwinia carotovora. Plant Mol Biol 59:
771–780.
130. Afroz, A., Chaudhry, Z., Rashid, U., Ali, G.M., Nazir, F., Iqbal, J., Khan, M.R. (2011). Enhanced resistance against bacterial wilt in
transgenic tomato (Lycopersicon esculentum) lines expressing the Xa21 gene. Plant Cell Tiss Organ Cult 104: 227–237.
131. Mendes, B.M.J., Cardoso, S.C., Boscariol-Camargo, R.L., Cruz, R.B., Mourão, F.A.A., Filho, A.B. (2010). Reduction in susceptibility to
Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis expressing the rice Xa21 gene. Plant Pathol 59: 68–75.
132. Urwin, P.E., Atkinson, H.J., Waller, D.A., McPherson, M.J. (1995). Engineered oryzacystatin-I expressed in transgenic hairy roots
confers resistance to Globodera Pallida. Plant J 8: 121–131.
133. Urwin, P.E., Lilley, C.J., McPherson, M.J., Atkinson, H.J. (1997). Resistance to both cyst and root-knot nematodes conferred by
transgenic Arabidopsis expressing a modified plant cystatin. Plant J 12: 455–461.
134. Urwin, P.E., Levesley, A., McPherson, M.J., Atkinson, H.J. (2000). Transgenic resistance to the nematode Rotylenchulus reniformis
conferred by Arabidopsis thaliana plants expressing proteinase inhibitors. Mol Breeding 6: 257–264.
135. Vain, P., Worland, B., Clarke, M.C., Richard, G., Beavis, M., Liu, H., Kohli, A., Leech, M., Snape, J., Christou, P., Atkinson, H. (1998).
Expression of an engineered cysteine proteinase inhibitor (Oryzacystatin-I Delta D86) for nematode resistance in transgenic rice
plants. Theor Appl Genet 96: 266–271.
136. Urwin, P.E., Troth, K.M., Zubko, E.I., Atkinson, H.J. (2001). Effective transgenic resistance to Globodera pallida in potato field trials.
Mol Breeding 8: 95–101.
137. Liu, B., Hibbard, J.K., Urwin, P.E., Atkinson, H.J. (2005). The production of synthetic chemodisruptive peptides in planta disrupts the
establishment of cyst nematodes. Plant Biotechnol J 3: 487–496.
138. Winter, M.D., McPherson, M.J., Atkinson, H.J. (2002). Neuronal uptake of pesticides disrupts chemosensory cells of nematodes.
Parasitology 125: 561–565.
139. Mesquita, A.L.M., Alves, E.J., Caldas, R.C. (1984). Resistance of banana cultivars to Cosmopolites sordidus (Germar, 1824).
Fruits 39: 287–290.
140. Ortiz, R., Vuylsteke, D., Dumpe, B., Ferris, R.S.B. (1995). Banana weevil resistance and corm hardness in Musa germplasm.
Euphytica 86: 95–102.
141. van den Berg, H., Faulks, R., Granado, H.F., Hirschberg, J., Olmedilla, B., Sandmann, G., Southon, S., Stahl, W. (2000). The potential
for the improvement of carotenoid levels in foods and the likely systemic effects.
Journal of the Science of Food and Agriculture 80: 880-912.
142. Davey, M.W., Van den Bergh, I., Markham, R., Swennen, R., Keulemans, J. (2009). Genetic variability in Musa fruit provitamin A
carotenoids, lutein and mineral micronutrient contents. Food Chem 115: 806–813.
143. Ekesa, B., Mirroir, C., Blomme, G., Van den Bergh, I., Davey, M.W. (2013). Retention of Provitamin A Carotenoids during Postharvest
Ripening and Processing of Three Popular Musa Cultivars in South-Western Uganda. International Ishs-Promusa Symposium on
Bananas and Plantains: Towards Sustainable Global Production and Improved Use 986: 319–330.
144. Mlalazi, B., Welsch, R., Namanya, P., Khanna, H., Geijskes, R.J., Harrison, M.D., Harding, R., Dale, J.L., Bateson, M. (2012). Isolation
and functional characterisation of banana phytoene synthase genes as potential cisgenes. Planta 236: 1585–1598.
145. Davey, M.W., Stals, E., Ngoh-Newilah, G., Tomekpe, K., Lusty, C., Markham, R., Swennen, R., Keulemans, J. (2007). Sampling
strategies and variability in fruit pulp micronutrient contents of West and Central African bananas and plantains (Musa species).
J Agr Food Chem 55: 2633–2644.
146. Ekesa, B., Nabuuma, D., Blomme, G., Van den Bergh, I. (2015). Provitamin A carotenoid content of unripe and ripe banana
cultivars for potential adoption in eastern Africa. J Food Compos Anal 43: 1–6.
147. Waltz, E. (2014). Vitamin A Super Banana in human trials. Nat Biotechnol 32: 857.
Bananas, the green gold of the South
55
IPBO (International Plant Biotechnologie Outreach), dat deel uitmaakt van het VIB, werd in 2000 opgericht door Prof.
Em. Prof. Marc Van Montagu en de Universiteit Gent, met steun van de Vlaamse Overheid en de Seghal Foundation.
De missie van IPBO is het bevorderen van kennis en technologietransfer in het domein van de plantenbiotechnologie
naar ontwikkelingslanden, met de focus op groene en duurzame landbouw en agro-industrie. Om dit te bewerkstelligen
richt IPBO zich op communicatie en training in plantenveredeling, groene biotechnologie en gerelateerde bioveiligheid
en op netwerking en projectontwikkeling om samenwerkingsverbanden tussen ontwikkelingslanden en Vlaanderen te
katalyseren. Meer info op www.ipbo.vib-ugent.be
Basisonderzoek in de levenswetenschappen, dat is de kernactiviteit van VIB. Enerzijds de grenzen verleggen van
wat we weten over moleculaire mechanismen, hoe deze mechanismen levende wezens zoals mense, dieren, planten
en microorganismen regelen, en anderzijds zorgen voor tasbare resultaten die bijdragen aan een betere samenleving.
Gestoeld op het parnerschap van vijf Vlaamse Universiteiten – UGent, KULeuven, Universiteit Antwerpen, Vrije
Universiteit Brussel en Universiteit Hasselt – en een stevig investeringsprogramma, bundelt VIB de expertise van 75
onderzoeksgroepen in één instituut. Het techtransfer-team van VIB vertaalt proactief biologische inzichten in nieuwe
economische activiteiten die op hun beurt weer kunnen leiden tot nieuwe producten, medicijnen e.d. die kunnen gebruikt
worden in de geneestkunde, landbouw en tal van andere toepassingen. VIB neemt ook actief deel aan het publieke
debat over biotechnologie door het ontwikkelen en verspreiden van een breed scala aan wetenschappelijk onderbouwde
informatie over alle aspecten van de biotechnologie. Meer info op www.vib.be
De Universiteit Gent (UGent), is met meer dan 41 000 studenten één van de grootste universiteiten van het Nederlandse
taalgebied. Het opleidingsaanbod omvat vrijwel alle academische opleidingen die in Vlaanderen worden ingericht.
De UGent profileert zich als een open, sociaal geëngageerde en pluralistische universiteit in een internationaal perspectief.
Meer info op www.UGent.be.
Het Laboratorium voor Tropische Plantenteelt maakt deel uit van de Faculteit Bio-ingenieurswetenschappen aan de
KU Leuven. Het wordt geleid door Prof. Dr. Rony Swennen en telt ongeveer 40 onderzoekers. Het onderzoek richt zich op
de productie van tropische gewassen, voornamelijk bananen, met verbeterde eigenschappen door zowel technieken uit
de klassieke veredeling, moleculaire plantenbiotechnologie, genoomanalyse en fenotypering met elkaar te combineren.
Het ultieme doel bestaat erin de levenskwaliteit van de kleine boeren te verbeteren door middel van duurzame landbouw.
Het laboratorium levert daarnaast een actieve bijdrage tot de bescherming van de biodiversiteit, beheert de grootste
bananencollectie (zowel in vitro als diepgevroren) en is de basis van “Bioversity International” in België.
NARO (National Agricultural Research Organization) heeft meer dan 25 jaar ervaring in het ontwikkelen van technologiën
om de productie van bananen in Oost en Centraal Afrika te verhogen. In deze gebieden ervaren de producenten
immers veel problemen door ziekten en andere bedreigingen. Het doel van NARI is om ziekte-resistente bananen en
bananen met verhoogde opbrengsten te ontwikkelen. Daarnaast ontwikkelde NARO via veredeling en biotechnologie
ook hooglandbananen met een verhoogde opbrengst, meer proivitamine A en resistentie tegen ziektes, nematoden en
snuitkevers. NARO ontwikkelde samen met IITA 27 EAHB hybriden. Twee van deze hybriden werden in 2010 op de markt
gebracht en worden door meer dan 15% van de bananenboeren in Oeganda geteeld.
IITA (International Institute of Tropical Agriculture), opgericht in 1967, is een van de toonaangevende instituten die
onderzoek verichten om honger, ondervoeding en armoede tegen te gaan, in het bijzonder in Sub-Sahara Afrika.
IITA werkt met verschillende onderzoekspartners samen om gewas kwaliteit en productiviteit te verhogen, risico’s voor
teler en consument te verlagen en een duurzame welvaart in de landbouw te ontwikkelen.
Het laboratorium voor Plantengenetica, veredeling en biotechnologie is gevestigd aan de Universiteit van Kisangani
(UNIKIS) in de Democratische Republiek Congo. Dit laboratorium werd opgericht en wordt nog steeds geleid door
Professor Benoit Dhed’a Djailo. De doelstellingen van dit laboratorium zijn de karakterisatie, het behoud, de selectie en
de verspreiding van gezonde bananenplanten. Sinds 2005 werden meer dan 100 verschillende soorten bakbananen
verzameld in het DR Congo, werden deze bewaard in de collectie en 97 hiervan werden reeds morfologisch
gekarakteriseerd.
UNIDO (United Nations Industrial Development Organization) heeft als doel armoede uit de wereld te helpen door
middel van inclusieve en duurzame industriële ontwikkeling (ISID). UNIDO pleit dat ISID de belangrijkste drijfveer is voor
de succesvolle integratie van de economische, sociale en milieuaspecten, die nodig zijn om duurzame ontwikkeling in het
belang van onze toekomstige generaties te bekomen.
IPBO
Technologiepark 3, 9052 Gent, België
Tel: + 32 9 264 87 27 - [email protected] - www.ipbo.vib-ugent.be
V.U. Sylvie De Buck (IPBO/VIB) en Rony Swennen (KULeuven) – D/2016/12.267/3