2016/2017
FPMO
De Kok
Fotosynthese: licht energie + 6CO2 + 6H2O = C6H12O6 + 6O2 (suikers
en zuurstof)
Net primary production = carbon fixatie per m2 per jaar
In praktijk is fotosynthese complexer: zie plaatje.
Fotosynthese vindt plaats in de chloroplast in het blad (zie plaatje voor
doorsnede blad). In het mesophyl zitten de meeste chloroplasten
(pallisade en spons). CO2 opname vindt plaats door de
stomata (onderkant van het blad). Deze kan de plant
reguleren. Een chloroplast is een structuur op zichzelf, bezit
alles wat een normale plantencel ook bevat + speciale
organellen. Stromafase is het cytoplasma van de
chloroplast. In de thylakoiden vindt de lichtabsorptie plaats.
Zetmeelkorrels kunnen voorkomen, zijn producten van
fotosynthese (opslag van suikers).
In bepaalde bacteria kan ook fotosynthese voorkomen,
maar deze gebruiken geen water maar H2S. Hier komt geen zuurstof uit maar zwavel (en water). De
zuurstof die uit fotosynthese komt komt dus niet uit de CO2 maar uit het water. Zie dia 18.
Fotosynthese in een chloroplast heeft twee fases, een licht- en donkerreactie. In de lichtreactie
worden energetisch rijke verbindingen gemaakt en wordt er ATP en NADPH aangemaakt. De
donkerreactie is niet afhankelijk van licht. Bij lichtreactie wordt licht geabsorbeerd en wordt energie
gemaakt en NADP wordt NADPH en ADP wordt ATP. Bij donkerreactie wordt de energie beruikt in de
Calvin cyclus om suiker, water en weer NADP en ADP te maken.
Energie kan door een redox reactie worden gemaakt (van NADP naar NADPH). Een andere
mogelijkheid is vormen van ATP. Lichtenergie zorgt ervoor dat ADP ATP wordt (fosfolylering reactie).
Lichtreactie heeft 3 stappen:
- Lichtabsorptie en excitatie van pigmenten
- Pigmenten worden overgegeven aan elektronen transport
- Chemiosmose (ATP synthese)
o Ophoping van H+ (protonen) in de chloroplast waardoor ATP wordt gegenereerd uit
ADP + P (fotofosfolylering).
Deze stappen vinden plaats in thylakoide membraan.
De zon geeft veel meer energie dan zichtbaar is.
Je hebt twee soorten chlorofylen (A en B). Verschillen in absorptie. Heel klein verschil in structuur.
Chlorofylen zijn geladen kop en lipide staart. Hierdoor vormen membranen (water afstotend en
aantrekkend). De staarten zijn waterafstotend.
Je hebt ook carotenoiden. Deze lijken op de lipiden. Je hebt 3 verschillende soorten. Caretenoiden
absorberen licht met een lagere golflengte (blauw licht) dan chlorofylen. B is net wat lager qua
golflengte dan A (beide rood-oranje licht). Als je dus een combinatie hebt van chlorofyl A en B is de
plant in staat meer golflengten licht te absorberen. Planten zijn groen omdat dat de golflengte is die
ze niet absorberen. In de herfst verkleuren planten door twee oorzaken, snel afgebroken
caretenoiden waarna andere pigmenten omhoog komen (die niet groen zijn). Deze andere
pigmenten zijn meestal wateroplosbaar en door het hele blad aanwezig (niet alleen in de
chloroplast).
Structuur van een chloroplast goed kennen (kunnen tekenen):
2016/2017
Belangrijke reacties in de lichtreactie:
Fotosynthese elektronen transport:
- Lichtreactie vindt plaats in thylokoide membraan.
- Er wordt NADPH en ATP gevormd wat later wordt gebruikt in de Calvin cyclus.
- Als er licht wordt geabsorbeerd door een pigment wordt een elektron aangeslagen.
- Hij kan dan het elektron doorgeven of het afgeven als warmte. Als licht wordt geabsorbeerd
kan een deel van de energie terugvallen als warmte of fluorescentie (licht afgeven).
- Een plant heeft twee fotosystemen (I en II). Ze lijken op elkaar maar zijn net wat anders. De
ene kan eerder absorberen dan de ander (II meer dan I). De chlorofyl ratio (A, B) verschilt ook
iets per fotosysteem. Beide systemen zijn nodig om zo effectief mogelijk licht om te zetten in
energie. Het licht gaat eerst in II, daarna in I.
o Een elektron komt uit water en
wordt geactiveerd door licht en
verhoogd in energie niveau (systeem
II). Deze wordt dan weer wat
afgebroken in energie waar ATP bij
vrijkomt en gaat in systeem I. Hier
wordt
het
elektron
weer
aangeslagen door licht waardoor het
elektron naar nog een hoger
energieniveau gaat waarna het weer
vervalt en er NADPH ontstaat (door
binding NADP+ en 2H+). Deze twee
vormen van energie gaan daarna de
Calvin cyclus in.
o Als 1 van de twee fotosystemen zou missen zou er alleen maar ATP ontstaan, wat
niet genoeg is.
o De systemen drijven in het thylakoide membraan.
o Aan de binnenkant van het membraan krijg je een ophoping van H+. Hierdoor werkt
het ATP synthase complex (fosfolylering van ADP naar ATP). Je hebt hier dus een
lokale verzuring.
o Uit de lichtreactie: 12NADP+ + 12 H2O + 18 ADP + 18 Pi 12 NADPH + 12H+ + 18
ATP + 6O2. Hierbij zijn 72 fotonen nodig. Dit is precies genoeg energie om 1 suiker te
maken.
o In de chloroplast wordt eigenlijk altijd de gefosfolyseerde vorm van NAD gebruikt
(NADP+).
Donkerreactie (Calvin cyclus):
- 6CO2 + 12NADPH + 12H+ + 18ATP
C6H12O6 + 12 NADP+ + 6H2O + 18
ADP + 18Pi.
- Calvin cyclus vindt plaats in het
cytoplasma van een chloroplast.
- Met de huidmondjes kan de plant de
zuurstof opname en waterhuishouding
reguleren. Deze worden gereguleerd
door de sluitcellen met turgor. Door
veel suikers in de sluitcellen te pompen
nemen deze veel water op waardoor je
een hoge osmotische druk hebt en de
huidmondjes open gaan staan.
2016/2017
-
-
De regulatie van huidmondjes in gematigde gebieden zijn niet zo van belang, alleen s nachts
gaan ze dicht. In extreme gebieden zijn ze echter sterk van belang (tegen uitdroging etc.).
Calvin Cyclus:
o fase 1: Koolstof fixatie. Daadwerkelijk vastleggen van CO2. CO2 bindt aan molecuul,
je krijgt een C6 suiker. RuBisCo splitst dit in 2 C3 moleculen.
RuBP (C5) bindt met CO2, krijg je een C6 molecuul, wordt door rubisco
omgezet in 2 C3 moleculen. (namen C6 en C3 en rubisco kennen!!).
o Fase 2: reductie. ATP en NADPH voor nodig. Substraat voor suikermoleculen wordt
gemaakt.
Je zet een glyceraat om naar een aldehyde dmv ATP en NADPH. Hierbij komt
glucose vrij. Wordt ook weer voor een deel gebruikt voor fase 3.
o Fase 3: Regeneratie of ribulose. Er ontstaat weer een C5 molecuul.
Je moet de fases en de substraten kennen.
Je hebt 6 CO2 moleculen, 6 watermoleculen, 18 ATP en 12 NADPH moleculen nodig voor 1
suiker molecuul (glucose).
Fotosynthese is licht gelimiteerd tot een bepaalde hoogte, daarna CO2 gelimiteerd. Bij een hele lage
licht absorbtie is de CO2 uptake meer dan de CO2 ‘evolution’.
C4 planten gaan langzamer in partiële CO2 druk omhoog, en
sneller in CO2 assimilatie, maar loopt dan zwakker af dan C3
planten. C4 planten gaan door de 0, C3 planten niet. Dit komt
door fotorespiratie.
Fotorespiratie = als zuurstof wordt gebruikt als
elektronenacceptor voor fotosynthese, ipv. CO2. RuBisCo
(ribulose bifosfaat carboxylase) doet dit onder hoge
temperaturen. Dit is ongunstig voor de plant, omdat je dan een C3 en een C2 molecuul krijgt.
Hierdoor is de Calvin cyclus dus half zo voordelig. De plant kan het C2 molecuul gebruiken om glycine
en serine te vormen. Uiteindelijk wordt er dan weer glyceraat gemaakt, waarna er weer RuBP wordt
gevormd en het weer terug kan in de CC. Hierbij komt CO2 vrij (vandaar dat het respiratie heet) en hij
gebruikt een extra ATP. In gematigde gebieden wordt ongeveer 20% van de Rubisco ‘verspild’ aan
fotorespiratie. Rubisco is waarschijnlijk geëvolueerd toen er nog weinig zuurstof in de atmosfeer was.
C3 planten gebruiken RuBisCo om 2 C3 verbindingen om te zetten in de
Calvin cyclus.
C4 planten vermijden fotorespiratie. Een belangrijke C4 plant is bijvoorbeeld
maïs. De morfologie verschilt van C3. Dit zit voornamelijk in het mesofyl. In
een C3 plant zit veel lucht. Een C4 plant heeft een heel compact
sponzenparengym. De vaatbundelcellen bij C4 planten zijn afgeschermd en
vergroot, en er zitten chloroplasten in (!), rondom de vaatbundels. C4
gebruiken PEPco ipv Rubisco. PEPco (PEP-carboxylase) zit in het mesofyl en
fixeert het CO2. Hierbij wordt een C4 verbinding gevormd (oxaalzuur). Dit
wordt omgezet naar malaat. Dit malaat wordt getransporteerd naar de
vaatbundelcellen. Hier wordt het vastgelegd en komt er CO2 vrij, door
middel van Rubisco. De Rubisco concentratie in deze cellen is heel hoog.
Nadat er CO2 vrij komt ontstaat er weer een C3 molecuul (PEP). Het
voordeel van dit mechanisme is dat de O2 heel slecht bij de
vaatbundelcellen kan komen. Hierdoor is de O2 concentratie in deze cellen
heel laag. Hierdoor vindt er (nagenoeg) geen fotorespiratie plaats. Ook
kunnen de producten die worden gevormd heel snel aan het floëem
afgegeven worden en worden getransporteerd.
2016/2017
De affiniteit van CO2 van PEPco is veel hoger dan die voor Rubisco. De affiniteit voor zuurstof van
Rubisco is veel lager dan die van CO2 (maar zuurstof komt ook veel meer voor).
Sommige planten hebben geen ruimtelijke verdeling (zoals C4), maar
een dag en nacht verdeling CAM planten. Deze komen vooral voor in
hele extreme omstandigheden (woestijnen). Overdag hebben ze hun
huidmondjes helemaal dicht en alleen in de nacht staan ze open, omdat
ze anders te veel water verliezen. Het probleem is dat ze dan overdag
geen toevoer van CO2 hebben. Daarom zijn ze in staat om s nachts CO2
te fixeren. S nachts staan mondjes open, CO2 wordt gefixeerd door
PEPco, wordt malaat gevormd, malaat gaat naar de vacuole. Overdag
wordt het malaat afgegeven uit de vacuole, wordt gedecarboxileerd,
komt CO2 vrij, wordt gebruikt door rubisco en gaat de Calvin cyclus in.
Eigenlijk hetzelfde als C4 dus, maar hier vindt het plaats in dezelfde mesofylcellen en is de scheiding
tussen nacht en dag. De meeste CAM planten gebruiken dit mechanisme niet altijd, alleen als het
echt nodig is. In de winter gaan ze terug naar een C3 (!) metabolisme.
C3 is het minst efficient met water, CAM het meest efficient. C4 planten komen meer voor in warme
gebieden en dichter bij de evenaar.
Fotosynthese
Zetmeelkorrels hebben dezelfde oorsprong als een chloroplast, komen uit een
amyloplast. Ook in een chloroplast zitten zetmeelkorrels. Producten die niet
meteen worden gebruikt uit fotosynthese kunnen worden opgeslagen als
zetmeelkorrels. Dit kan soms teveel zijn waardoor de plant schade ondervindt (bij
een te hoge CO2 concentratie), meestal aan de chloroplast zelf. In de wortels
zitten amyloplasten. Statolithen bevatten ook zetmeel en hebben een belangrijke
rol bij het naar beneden groeien van de wortel. Het rode van een tomaat komt
doordat de chloroplasten om worden gezet in chromoplasten. Suikers worden in
de plant opgeslagen als sucrose. Deze worden getransporteerd door het floëem,
het xyleem (houtvaten, dood) is belangrijk bij opname van mineralen en transport van water
(beneden naar boven). Suikers gaan via het floëem van boven naar beneden en beneden naar boven.
Suikers gaan eerst door de companion cells in het floëem. Dit is een actief proces met de proton
force. Suikertransport is dus een actief proces. De suikers worden geladen in de floeem cellen. Als
deze concentratie hoog is neemt de plant zoveel mogelijk water op (turgor) uit de xyleemvaten.
Hierdoor krijg je een hoge osmotische druk. Zo krijg je verschillende turgor drukken (op plekken waar
suiker nodig is laag, waar suiker niet nodig is hoog) waardoor je een stroom krijgt. Dit kan richting de
wortels zijn maar ook richting de toppen (voor groei). Als de druk in de wortels ook hoog is krijg je
een opslag in de chloroplasten/cellen van suikers in bijvoorbeeld zetmeelkorrels. Suikergebruik
wordt dus beïnvloed door de sink demand.
De Calvin cyclus wordt niet alleen gebruikt voor de aanmaak van suikers, maar ook voor de synthese
van andere producten zoals lipiden.
2016/2017
De ademhaling van de plant
Hierbij komt juist CO2 vrij. Het is geen echte ademhaling, maar ze gebruiken zuurstof en er komt CO2
vrij. Hierdoor komt toch heel veel koolstof die wordt opgenomen door fotosynthese weer vrij in de
atmosfeer.
Een plant heeft altijd een vaste maintenance respiration waar de growth respiration nog boven op
komt. Deze bepalen samen de kosten voor de groei.
De ademhaling vindt plaats in de mitochondria. Het
gaat van het cytoplasma naar de chloroplast, waarna
het naar het mitochrondia gaat (??). Voor de afbraak
van 1 suiker molecuul heb je 6 zuurstof moleculen
nodig. Bij de afbraak hiervan komt 36 ATP vrij, 6CO2
en 6H2O.
Glucose gaat in de cel, er vindt glycolyse plaats waarbij
ATP en NADH vrij komt. Dit wordt omgezet in
pyruvaat, waarna dit wordt omgezet in lactaat en
acetyl-CoA en NADH. Acetyl-CoA gaat de Krebs cyclus
(citroenzuurcyclus) in waarbij weer ATP en NADH vrij
komt, en CO2 gaat weer uit de cel. NADH wordt
gebruikt in het elektronen transport systeem, wat
weer ATP oplevert. Zuurstof gaat de cel en het
mitochondrium in en wordt H2O. Glycolyse is
belangrijk, zie plaatje.
Pyruvaat wordt met zuurstof anders omgezet dan
zonder zuurstof. Zonder zuurstof wordt het
acetaldehyde waarna het ethanol wordt. Met zuurstof
wordt het acetyl-CoA of lactaat (voor spieren). AcetylCoA gaat de krebs cyclus/citroenzuurscyclus in waarbij
ATP, NADH en 2CO2 vrij komt.
Actieve fosfolilering (chemiosmose): NADH en FADH2
wordt omgezet in ATP. Dit gaat via een
elektrontransportketen.
Als een plant in een anaerobe omgeving komt kan het aerenchym gaan aanmaken, een soort
parenchym maar dan met veel lucht tussen de cellen. Hierdoor kan het ook respiratie doen met
andere stoffen dan zuurstof.
Suikers zijn niet de enige energiebron voor planten. De skunk cabbage heeft de meest precieze
temperatuur regeling in planten die we tot nu toe hebben gevonden. Hij kan zelfs sneeuw laten
smelten. Andere planten doen ook aan warmte regulatie, bijvoorbeeld de philodendron
solimoesense. Dit is een vorm van alternatieve respiratie: thermogenesis.
Minerale huishouding
Alleen vlinderbloemigen hebben een symbiose met wortelknolletjes.
Andere stoffen dan CO2 worden voornamelijk in de wortels opgenomen. De belangrijkste zijn zwavel,
fosfor en stikstof. Er zijn 13 essentiële elementen voor een plant. De metalen de voorkomen in een
plant zijn vaak afhankelijk aan de enzym omzetting, niet aan de groei oid. Hierom is er maar weinig
van nodig.
Hoe worden mineralen opgenomen? Dit gaat meestal met actief transport of facultair. Voor de
meeste mineralen zijn er specifieke transporters, vaak meer dan één. Er zijn twee manieren van
opnemen:
- Symplastische route: het mineraal gaat meteen door de membranen het cytoplasme in. Dit
heet symplastisch transport. Tussen de cellen is vrij transport mogelijk door het cytoplasma.
2016/2017
-
Apoplastische route: het mineraal gaat via de celwand naar de binnenkant van de wortel. Het
endodermis is een barrière door het bandje van casparie. Deze is impermeabel voor
voornamelijk mineralen. Hier moet het mineraal wel door het endodermis en het cytoplasma
(dus laatste stukje altijd symplastisch).
Hierna worden de mineralen afgegeven aan het xyleem. De drijvende kracht achter de opname van
mineralen is verdamping. Het xyleem vervoert de mineralen en water.
Stikstof en zwavel worden opgenomen in een geoxideerde vorm (kan niet anders). De plant kan
echter niks met SO4=(sulfaat) en NO3-(nitraat). Deze moeten dus eerst worden gereduceerd. De
reductie vindt voornamelijk plaats in de chloroplast. Sulfaat helemaal, nitraat wordt eerst omgezet in
nitriet en gaat dan naar de chloroplast. Er wordt ongeveer 15 keer zoveel nitraat gebruikt dan
sulfaat.
Reductie van nitraat:
NO3- + 10 H+ + 8 elektronen NH4+ + 3H2O
Reductie van sulfaat:
SO4 2- + ATP + 8 H+ + 8 elektronen S 2- + 4H2O + AMP + PPi
Nitraat is niet de enige stikstofbron die een plant kan opnemen. Een plant kan ook ammonium
(NH4+) opnemen. Deze kan hij meteen gebruiken in aminozuren. Ammonium is in te hoge
concentraties echter giftig voor een plant.
Vlinderbloemigen hebben een symbiose met bacteriën waardoor ze pure stikstof kunnen opnemen
(N2) (uit de lucht). Deze bacteriën zijn rhizobium. Ook kunnen sommige planten insecten opnemen
als bron voor stikstof.
Rhizobium bacteriën vormen knolletjes in de
wortels van vlinderbloemigen. Hiermee kunnen
planten atmosferische stikstof omzetten in
ammonium. Hierbij moet wel zuurstof aanwezig
zijn. Het opnemen van deze bacteriën is van de
plant een actief proces. Een wortelhaar krult om
de bacteriën heen, waarna je een infectiedraad
krijgt. Hierna krijgt de wortel een morfologische
verandering waarbij een afscherming plaats vindt
waarin de bacteriën aanwezig zijn. Hierna worden
de cellen van het epidermis verschoven
(inkapseling), waardoor er blaasjes ontstaan. In
deze knolletjes kan stikstoffixatie plaats vinden.
Er vindt dan nitrogenase plaats waardoor N2
wordt omgezet in ammonium (NH4+). Deze
reductie kan alleen maar plaatsvinden in aanwezigheid van
zuurstof. Daarom maken de planten in de blaasjes
leghemoglobine aan, wat heel hoge affiniteit heeft voor
zuurstof. Hierdoor blijft de zuurstofconcentratie in de blaasjes
hoog. Dit is een heel effectief systeem voor het opnemen van
stikstof. In de herfst komt deze stikstof weer vrij. Je kan ook
vlinderbloemigen tussen andere gewassen planten zodat je
meer stikstof in de bodem krijgt. Hoe minder nitraat in het
xyleemsap en hoe meer aminozuren of afgeleiden hiervan, hoe
groter de kans is dat de plant alleen maar atmosferische stikstof
gebruikt.
Planten kunnen ook schimmels gebruiken voor de opname van nutriënten. Ongeveer 80% gaan
symbiose aan met schimmels. Deze schimmels zijn mycorrhiza. Je hebt twee vormen: arbusculaire
mycorrhiza en ectomycorrhiza. De schimmels gaan in de wortel zelf zitten waardoor het oppervlakte
van de wortels enorm wordt vergroot. De schimmel krijgt dan fotosynthese producten voor terug.
2016/2017
Arbusculair (endo) gaat wat meer in de cellen zelf zitten, ecto zit meer aan de buitenkant van de
wortels en tussen de cellen.
2016/2017
Dijkhuizen
Microorganisms, diversity and cell structure
1. Beter begrijpen van levende organismen (Wat is leven?)
2. Die kennis toepassen voor het voordeel van mensen
a. Belangrijke rollen in medicijnen, landbouw en industrie.
Onder de prokaryoten verstaan we bacteriën en archaea, onder de eukaryoten onderscheiden we
eukarya. Dit is al het leven. Het grote verschil is dat de eukaryoot een celkern heeft. Deze heeft
sowieso meer celorganen. De mitochondria waren ooit bacteriën. Zelfde verhaal met chloroplasten.
Archaea zijn dichter verwant met eukarya dan met bacteria (latere afsplitsing). Deze boom is
gebaseerd op 16S ribosomaal DNA sequencies.
Archaea worden meer gevonden in extreme milieu’s.
In de pens van koeien zitten veel bacteriën die cellulose afbreken. Wij (de mens) kunnen cellulose
niet goed afbreken. Het gras wordt cellulose, wordt glucose, wordt microbiele fermentatie, wat
opsplitst in vetzuren en CO2 + CH4 (afvalproducten).
Wortelknolletjes helpen planten met het opnemen van N2 door dat om te zetten samen met H naar
NH3, wat de plant wel kan opnemen. In ruil daarvoor krijgen de bacteriën voedingsstoffen en
mineralen.
In ons lichaam competeren goede bacterien met pathogenen voor plaatsen en resources. Daarom
niet te veel wassen. Veel bacteriën in de maag en darm, hoe hoger de pH hoe meer bacteriën, en hoe
dichter bij de endeldarm.
Kleine cellen hebben meer contactoppervlak met het milieu er om heen waardoor ze beter
voedingsstoffen op kunnen nemen. Daardoor kunnen bijvoorbeeld coccen heel snel groeien. Hierbij
is het cytoplasma membraan van groot belang. Deze bestaat uit een dubbele laag van fosfolipiden.
De koppen bestaan uit hydrofiele glycerol met een fosfaat en/of ethanolamine staart. De staarten
zijn hydrofobe vetzuren. In deze laag zitten veel eiwitten verankerd, zoals bijvoorbeeld integrale
membraan eiwitten die door het hele membraan zitten. Dit zijn meestal transporteiwitten. Sommige
steken alleen aan de buitenkant er uit, dit zijn meestal receptoreiwitten. Drie functies van het
membraan:
1. barriere
2. eiwitanker
3. energie conservatie door een protonen gradiënt.
Verschillende vormen van actief transport (dia 30):
- Simpel transport: wordt gedreven door de proton motive
force (H+ en OH-).
- Groep translocatie: tijdens het transport wordt het
molecuul gefosfolileerd.
- ABC transporter: drie eiwitten bij betrokken. Aan de
buitenkant is een bindingseiwit aanwezig met een hoge
affiniteit om het molecuul dat binnen komt te binden. Deze
ligt in het periplasma. In het membraan zit een transport
eiwit. Aan de binnenkant zit een ATP gebruikend eiwit.
Het peptidoglycan is bij gram-positieve bacteriën heel groot.
Hierdoor kunnen we deze bacteriën marken met jodium waardoor ze paars kleuren. Een g-neg
bacterie heeft een hele dunne laag, waardoor ze de jodium niet vasthouden en ze rood kleuren. Ook
hebben deze bacterien aan het buiten membraan polysachiriden en hebben vaak een toxisch
membraan. Gram-negatieve bacteriën hebben vaak meer reliëf aan de buitenkant (te zien onder de
microscoop).
2016/2017
Lysozyme wordt vaak gebruikt om membranen op te breken doordat ze de beta 1,4 binding
verbreken tussen de acetylzuren in.
Veel bacteriën kunnen kapsels hebben. Dat zijn over het algemeen polysacchariden. Deze kunnen
een stevige laag om de bacterie heen vormen om zich tegen toxische milieus te beschermen (zoals
een zuurstofrijke omgeving). Ze kunnen ook die kapsels maken om zich te hechten aan oppervlakten
(biofilms).
Andere features zijn flagellen, waarmee ze zich kunnen voortbewegen. Deze zijn vaak langer dan de
bacterie zelf. Ook kunnen ze fimbriae hebben, waarmee ze zich kunnen vasthechten of DNA over
kunnen geven aan andere bacteriën. Ook kunnen bacteriën alkanoate aanmaken wanneer ze heel
veel koolstof hebben en te weinig stikstof. Dan kunnen ze de koolstof opslaan. Nog andere vormen
die je ziet in cellen is dat ze zwavel opstaan. Deze gebruiken sulfide (H2S) als energiebron en ze
maken daar zwavel van.
De groene aanslag in meren is vaak blauwalg. Door zonlicht kan de cyanobacterie zijn blaasjes vullen
en naar het oppervlakte drijven om meer zonlicht te vangen.
Endosporen zijn gedifferienteerde cellen die resistent zijn tegen bijvoorbeeld hitte. Dit kan je zien als
een soort dormant stage. Er blijft een spore achter die overleeft en wacht tot de situaties beter zijn.
Ze zijn alleen in gram-positieve bacteriën aanwezig. Het sporen maken gebeurt alleen aan het einde
van de groei. Calcium is erg belangrijk voor sporevorming.
Flagellen kunnen polair zijn en over het hele oppervlakte verspreid zijn, of juist aan één kant. De
motor hiervoor in de negatieve cel werkt op een proton moter force. Flagellen komen ook voor in de
gram-positieve bacterie. Door de flagellen kunnen bacteriën naar een plek gaan waar ze het liefst
zijn, denk aan attractants en repellents.
2016/2017
Microbiële groei
Celdeling: Cell elongation septum (tussenwand) formation completion of septum; formation of
walls; cell separation
4 stadia groei bacteriën: lag fase (aanpassingsperiode), exponentiële groei, stationair (zelfde
niveau, lichte groei, soms sporen maken), dood
(voedingsbodem is op). De troebelheid is te meten in
de stationaire fase.
- Lag fase: periode na enten en voor groei
- Exponentiele fase: cellen het gezondst, hard
groeien
- Stationaire fase: evenveel groei als dood.
Bodem raakt op of te veel afval
- Dood fase: de cellen gaan in lysus.
Hoe meet je het aantal cellen?
- Telkamer (onder de microscoop). Hierbij kijk je niet of ze levend of dood zijn.
- Agarplaat methode. Cultuur uitspreiden over een agarplaat, incuberen en daarna kolonies
tellen. Andere methode is de cultuur eerst op de plaat, daarna de agaar en dan incuberen.
Dan kan je zien welke kolonies op de agar zitten en welke in/onder de agar. Hieraan kan je
zien welke anaeroob en aeroob zijn. Hierbij tel je alleen levende cellen.
- Verdunningsreeks in buisjes. Als je cultuur te groot is en je niet meer kan tellen. Zo verdun je
steeds 10 keer (per buisje).
- Spectrofotometer. Deze meet de optische dichtheid. Je meet hoeveel licht er wordt
gereflecteerd, hoe meer cellen, hoe minder licht er door heen gaat.
Bij lagere temperaturen, lagere groeisnelheid.
- Prokaryoten kunnen bij hogere temperaturen groeien dan eukaryoten.
- Organismen met de hoogste temperatuur optima zijn meestal Archaea.
- Niet fototrofe organismen kunnen bij hogere temperaturen groeien dan fototrofe
organismen.
Sommige cellen hebben vanbinnen een mechanisme om zout tegen te gaan, een soort ‘tegenenzym’
(compatible solutes), zodat de osmotische waarde in de cel hoog blijft en het zout buiten de cel blijft.
Deze enzymen hebben verder geen invloed op de cel en zijn dus neutraal.
2016/2017
Micro-organismen zijn onder te verdelen in aeroob en anaeroob. Onder deze groepen zijn nog
subgroepen, zoals aeroob obligaat (heeft zuurstof nodig), aeroob facultatief (niet nodig maar wel
beter), aeroob microearofilisch (heeft maar een beetje nodig, minder dan in de atmosfeer, maar
heeft het wel echt nodig), anaeroob aerotolerant (niet nodig, en groei niet beter als er geen zuurstof
aanwezig is), anaeroob obligaat (zuurstof is schadelijk of dodelijk). Al deze vormen hebben een
bepaald type metabolisme waarbij ze wel of geen
zuurstof nodig hebben. Voorbeelden hiervan zijn
aerobe
ademhaling,
vergisting/fermentatie,
anaerobe ademhaling (met nitraat). Deze moet je
kennen zodat je de buisjes kan benoemen:
a) obligaat aeroob
b) obligaat anaeroob
c) aeroob facultatief
d) aeroob microearofilisch
e) anaeroob aerotolerant
Lecture 5
Nutriënten, media, diversiteit en eenheid in bioenergetica
Macronutrienten: C (koolstof), N (stikstof, eiwitten, DNA, RNA), P (fosfaat), S (zwavel, aminozuren
(3/20)), K, Mg, Ca, (Na), (Fe) (en zuurstof voor de aeroben).
Micro-organimsen hebben ook micronutrienten nodig:
Drie verschillende typen media:
- defined medium: we kennen alle componenten
- complex medium: compositie is niet helemaal bekend in detail
- minimaal medium: alleen de hoognodige elementen zijn aanwezig
3 energie klasses van micro-organismen
- metabolisme
o de som van alle chemische reacties die plaatsvinden in een cel
- catabolisme
o energie vrijmakende metabolische reacties
2016/2017
-
groepen energie klassen
o chemorganotrofen
o chemolithotrofen
o fototrofen
o heterotrofen
o autotrofen
Energie van redox reacties wordt gebruikt in de synthese
van energierije verbindingen (bijv. ATP). Redox reacties
gebeuren in paren (twee halve reacties). De elektron
donor is de stof die wordt geoxideerd in een redox
reactie. De elektron acceptor is de stof die wordt gereduceerd in een
redox reactie. De reductie potentiaal (E0’) geeft aan hoe graag een
stof elektronen wil afstaan. Dit wordt uitgedrukt in volt. Onder
verschillende omstandigheden kunnen stoffen een donor of acceptor
zijn (redox koppel). De gereduceerde stof met een meer negatieve E0’
doneert elektronen aan de geoxideerde stof met een meer positieve
E0’. Dit is te zien in de redox toren. Hoe hoger een stof in de redox
toren, hoe eerder hij doneert. Hoe lager een stof in de toren, hoe
eerder hij accepteert. Hoe groter het verschil tussen de twee stoffen,
hoe meer energie er bij vrij komt en dus hoe ‘liever’ de reactie plaats
vindt.
Chemische energie die vrijkomt in een redox reactie wordt
voornamelijk opgeslagen in bepaalde gefosforyleerde verbindingen.
- ATP, de grootste energie currency
- Phospoenolpyruvaat
- Glucose 6-phosfaat
- Co-enzym A
Bij de glycolyse wordt glucose geconsumeerd, twee ATP’s gevormd en fermentatie producten
gevormd. Sommige van deze producten kunnen worden gebruikt voor menselijk gebruik.
Bij aerobe respiratie wordt O2 geoxideerd en gebruikt als elektronen acceptor. Uit deze reactie komt
meer ATP vrij dan uit fermentatie (is dus voordeliger). ATP wordt geproduceerd door de proton
motive force, die draait op elektronen transport.
Elektronen transport systemen zijn membraan geassocieerd. Ze mediëren het ‘vervoer’ van
elektronen en gebruiken de energie die
hier bij vrij komt om ATP te synthetiseren.
Veel redox enzymen zijn betrokken bij het
elektronen
transport
(bijv.
NADH
dehydrogenase, flavoproteinen, ijzerzwavel eiwitten, cytochromen). Het
elektronen
transport
systeem
is
georiënteerd in het cytoplasmisch
membraan zodat elektronen gescheiden
zijn van protonen. Elektronen dragers
liggen in het membraan op volgorde van
hun reductie potentiaal (E0’). De laatste
drager in deze keten doneert het
elektron/proton an de elektronen
acceptor.
Tijdens deze elektronen transfer worden
verschillende protonen vrijelaten naar buiten het membraan. Deze protonen komen van NADH en
het uit elkaar vallen van water. Dit resulteert in een generatie van een pH gradient en een
2016/2017
elektrochemische potentiaal door het membraan (de proton motive force). De binnenkant van het
membraan wordt elektrisch negatief en basisch. De buitenkant wordt positief en zuur.
ATP synthase (ATPase) is een complex dat de proton motive force omzet in ATP. Het bestaat uit twee
onderdelen:
- F1: veel eiwitten complex, aan de binnenkant van de cel maar buiten het membraan, naar
het cytoplasma gericht.
- F0: protonen conduting binnenmembraan
kanaal.
- Het is reversibel.
Citric acid cycle (CAC) (citroenzuurcyclus) is een manier
waarop pyruvaat compleet wordt geoxideerd:
- De eerste stappen zijn hetzelfde als in de
glycolyse (van glucose naar pyruvaat).
- Per glucose molecuul, komt er 6 CO2 vrij en
NADH en FADH wordt gegenereerd.
- Speelt een belangrijke rol in catabolisme en
biosynthese.
- Heeft een energetisch voordeel over aerobe
respiratie.
Lecture 6
Anaerobe respiratie
Bij anaerobe ademhaling wordt een andere elektronen acceptor gebruikt dan O2. Verder lijkt het
sterk op aerobe ademhaling, alleen komt er minder energie uit anaerobe ademhaling. Hoe veel
energie er uit de ademhaling komt kan worden bepaald door de reductie potentialen te vergelijken
(redox toren). Bij anaerobe ademhaling is stikstof vaak de elektronen acceptor. De producten van de
nitraat reductie (denitrificatie) zijn gassen. Denitrificatie is de grootste biologische bron van N2 in
gasvorm.
Denitrificatie: nitraat (NO3-) nitriet (NO2-) nitrische oxide (NO) (gas) nitrous oxide (N2O)
(gas) dinitrogen (N2) (gas).
De eerste stap, van nitraat naar nitriet heet nitraat reductie en gebeurd bijvoorbeeld in E. coli. De
hele denitrificatie gebeurd bijvoorbeeld in Pseudomonas stutzeri. Enzymen die meehelpen aan deze
reactie keten worden onderdrukt door zuurstof.
Anaerobe ademhaling kan ook gaan met zwavel, dit heet de
sulfaat reductie. Reductie van SO4 2- naar H2S gaat door
verschillende intermediates en heeft activatie van sulfaat door ATP
nodig. Je hebt twee soorten sulfaat reductie: dissimilatieve en
assimilatieve sulfaat reductie. Assimilatieve heeft een extra ATP
nodig en een NADPH. Gedurende sulfaat reductie leidt het
elektronen transport rectie tot een proton motive force formatie.
Dit zorgt voor ATP synthese door ATPase. Veel verschillende
moleculen kunnen de elektronen donor zijn in een sulfaat
reductie, bijvoorbeeld H2, organische moleculen, fosfiet. Sommige
sulfur reducerende bacteriën kunnen extra energie krijgen door
disproportionation van sulfur moleculen.
Acetogenen en methanogenen gebruiken CO2 als een elektronen
acceptor in anaerobe respiratie. H2 is de grootste elektronen
donor voor beide groepen organismen. Dit heet acetogenese. Het
verschil tussen methanogenese en acetogenese is: bij methanogenese is de elektronen acceptor 1
HCO3- en komt er CH4 + 3 H2O uit. Bij acetogenese gebruiken ze 2 HCO3- moleculen als elektronen
acceptor en komt er C2H3O2- en 4 H2O uit. Ook komt er uit methanogenese meer energie vrij.
2016/2017
Micro-ogranismen laten dus een grote verscheidenheid aan mechanismen voor energie generatie
zien, zoals:
- fermentatie
- aerobe respiratie
- anaerobe respiratie
- chemolithotrofie
- fototrofie
Werkcollege I
1. het membraan is opgebouwd uit fosfolipiden, eiwitten en polisachariden. De functies van de
poriën is dat er moleculen in en uit de cel kunnen, al zijn ze meestal selectief. Ze zitten in het
buitenmembraan. Het lipide-A heeft endotoxische eigenschappen. Hierdoor heeft het een
effect op bijvoorbeeld darmweefsel. Bij gram-positieve bacterie is een hele dikke celwand
peptidoglycanlaag, bij negatieve een hele dunne. Het jodium blijft hangen in die dikke
celwand, maar niet in de dunne.
2. Fosfolipiden-bi-laag, opgebouwd uit vetzuurketens en glycerolfosfaat, waar ook groepen
fosfaat en ethanolamine in kunnen zitten. 3 functies zijn permeabele barrière, eiwit anker,
energie conservatie.
3. Een flagel is opgebouwd uit eiwitten die in de vorm zitten van een lange draad, en hij heeft
bepaalde ringen (ook eiwitten) in de drie lagen van de gram negatieve cel. Ze komen ook
voor bij positieve cellen maar hebben dan minder ringen. De flagel wordt aangedraaid door
de proton motive force van motoreiwitten. Chemotaxis is de verandering van richting van
organismen als reactie op chemicaliën (denk aan buisjes in bakjes).
4. Koolstof, stikstof, zuurstof, waterstof (zwavel, fosfor). Stikstof komt als ammonium of nitraat
in de cel (of aminozuren). Koolstof, waterstof en zuurstof komen van de lucht. Fosfor zit in de
fosfolipidenbilaag, DNA en RNA en ATP. Een bacteriële cel bestaat voor 50% uit eiwitten.
Functies zijn transport over membranen, enzymen van glycolyse en citroenzuurcyclus,
eiwitten in de elektronen transportketen, ATPase.
5. E. coli zou het snelst groeien in het complexe medium, hier zitten al veel stoffen
(celcomponenten) in die hij dan zelf niet meer hoeft te maken. E. coli heeft altijd een
koolstofbron nodig.
6. H2 is de elektron donor. De elektronen acceptor is O2. Waarom is nitraat een betere
acceptor dan fumaraat? Omdat fumaraat redelijk dicht bij waterstof staat in de redoxtoren,
waardoor het reductie verschil minder groot is dan nitraat. Voorbeelden van redoxtoren in
dia 14 kennen. Waterstof heeft de beste reductiepotentiaal, want deze is sterker negatief.
Hierdoor komt er met de reactie met zuurstof meer energie vrij. NAD is een betere
elektronenacceptor dan H+, omdat hij lager in de toren staat.
7. Dit moet je kunnen tekenen. Zie dia 15 en 16.
8. Het is exponentiële groei omdat elke cel zich kan vermenigvuldigen. Een semilogaritmische
plot is het aantal cellen op een logaritmische schaal tegenover een tijdschaal, waardoor het
een lineaire plot wordt. Hier kan je bijvoorbeeld de groeisnelheid halen. Als je heel zorgvuldig
een cultuur oment in hetzelfde medium is er geen lag fase. Obligaat aeroob kan alleen
groeien met zuurstof, dus bovenin het buisje. Obligaat anaeroob groeit alleen onder in
(zonder zuurstof). Facultatief aeroob is meeste bovenaan, maar ook onderin. Microaerofiel is
gevoelig voor zuurstof maar heeft het wel een beetje nodig, hij groeit dus vlak onder de
zuurstof grens en een beetje in zuurstof. Aerotolerant groeit door het hele buisje.
Lecture 7
Stikstoffixatie en symbiose (bacteriële kant).
Stikstof heeft een heleboel oxidatie vormen. Als stikstof wil worden opgenomen dan is het bijna altijd
een reducerend proces (wordt opgenomen als ammonia, -3 elektronen). Stikstof fixatie is als N2 gas
wordt gebruikt als stikstof bron in de cellen. Hiervoor moet het eerst omgezet worden in NH4+: dit
2016/2017
heet nitrogenase. Deze reactie is echter zeer gevoelig voor
zuurstof, waardoor de reactie mis kan lopen. Er zijn
verschillende manieren van micro-organismen om dit te
voorkomen. Stikstoffixatie wordt maar door een beperkte
groep prokaryoten uitgevoerd (alleen de organismen met ??? in
het DNA). Zie plaatje voor een overzicht van organismen die
aan stikstoffixatie doen (zowel vrijlevende aeroben als
anaeroben en symbiotische organismen).
De symbiose van stikstoffixatie is een van de belangrijkste
symbioses die we kennen. Rhizobia zijn hiervan de meest
bekende bacteriën. Molecuul kennen.
Nitrogenase reactie: N2 N2H2 (komt H2 vrij en 4H er in)
2 H2N (2 H nodig) 2 NH3 (2 H nodig)
Bij deze reactie is 16 ATP nodig.
Sommige bacteriën maken een slijmlaag om zichzelf heen om
zich te beschermen tegen zuurstof (Azotobacter). Dit is een van
de methoden. Cyanobacteriën die in lange ketens leven hebben
speciale cellen. Deze cel voert alle stikstoffixatie uit. Dit is de
enige cel waarin geen fotosynthese plaatsvindt. Dit is de
heterocyst. In de overige vegetatieve cellen vindt wel
fotosynthese plaats. Deze voorzien de heterocyst wel met
bepaalde moleculen, zoals pyruvaat. Dit is nodig voor het
reduceren van stikstof. Het glutamine dat de heterocyst
aanmaakt wordt naar de buurcellen vervoert. De derde aanpassing is een hemoglobine achtig
molecuul (leghemoglobine) dat in staat is zuurstof weg te vangen. Dit geeft een rode kleur. Dit
gebeurt bijvoorbeeld in wortelknolletjes.
Het doen van nitrogenase kost erg veel energie. Dit indiceert dat het zeer gereguleerd wordt (als
ammonia niet nodig is wordt het ook niet aangemaakt). Alle enzymen en onderdelen die een cel
nodig heeft voor nitrogenase ligt in een cluster in het DNA. Als er bijvoorbeeld ammonia voorkomt in
het medium zal de cel de onderdelen en enzymen niet gaan aanmaken voor nitrogenase.
Door wortelknolletjes kunnen planten stikstof uit de lucht halen.
Lecture 8
Licht als energiebron
- Fototrofy: gebruikt licht als energiebron
o Fotofosforylatie: licht gemediëerde ATP synthese
o Fotoautotrofen: gebruiken ATP voor assimilatie van CO2 voor biosynthese
o Fotoheterotrofen: gebruiken ATP voor assimilatie van een organisch C molecuul voor
biosynthese.
Fototrofe organsimen gebruiken licht als energiebron.
Fotofosforylatie is ATP synthese dmv licht. Dit is wat anders
dan vanuit water. Fotoheterotrofen hebben geen calvin cyclus.
Deze gebruiken lichtenergie maar kunnen ze uitgaan van hun
organische molecuul (gebruiken geen CO2).
Het pigment dat Archaea bacteriën rood maakt is
bacteriorhodopsine. Een onderdeel hiervan (RetT) gaat van
trans naar cis, waardoor er zich een protonenpomp vormt.
Hierdoor wordt ATP gevormd. Dit noemen we geen
fotosynthese omdat er geen chlorofyl pigmenten aan mee
doen.
2016/2017
Fotosynthese is de conversie van licht energie naar chemische energie. Fototrofen doen aan
fotosynthese. De meeste fototrofen zijn autotrofen. Fotosynthese heeft licht gevoelige pigmenten
nodig, genamad chlorofyllen. Fotoautotrofie heeft ATP productie en CO2 reductie nodig.
Fotoheterotrofen gebruiken geen Calvin cyclus
voor de C productie, maar organische bronnen
(dit zijn meestal bacteriën). Je hebt oxygene
(cyanobacteriën) en anoxygene (paarse en
groene bacteriën) prokaryote fototrofen.
Bacteriën gebruiken een bacteriochlorofyl dat
erg veel lijkt op het planten chlorofyl. Het heeft
net een andere zijgroep. Chlorofylen hebben een
bepaalde ringstructuur met daarin een
magnesium molecuul. Ze vangen wel andere
golflengten dan planten (360, 805 en 870). Deze
zitten allemaal in chlorosomen om de rand van de bacterie (zie plaatje). Carotenoiden
worden altijd gevonden in fototrofe organismen. Het voorkomt fotooxidatie en ze
kunnen energie invangen en naar het reactiecentrum brengen.
Lecture 9
Chemolithotrofie
Een chemolithotroof organisme gebruikt anorganische chemische moleculen als
elektronen donor. Voorbeelden zijn H2S, H2, Fe2+ en NH3 (ammonia). Deze organismen
zijn meestal aeroob en begint de oxidatie met een anorganische elektronen donor. Het
gebruikt de elektronen transport keten en proton motive force en is autotroof: het
gebruikt CO2 als koolstofbron. Dit is volgens de redox toren echter minder succesvol qua
energie opbrengst.
H2 oxidatie
Anaerobe H2 oxidiserende bacteriën en archaea zijn bekend. Ze
katalyseren door hydrogenase. Door de calvin cyclus en hydrogenase
enzymen kan er chemolithotrofe groei plaatsvinden.
Vaak wordt zwavel ook gebruikt als elektronen donor. Voornamelijk H2S,
S0 en S2O3- worden veel gebruikt. Een product van zwavel oxidatie is H+,
waardoor de pH van het milieu omlaag gaat. Sox systemen oxideren
gereduceerde zwavel moleculen direct in sulfaat. De meeste organimsen
die aan zwavel oxidatie doen zijn aeroob, maar sommige kunnen nitraat als
een elektronen acceptor gebruiken. Elektronen uit het gereduceerde
zwavel komen bij het elektronen transport systeem. Vanaf hier worden ze
getransporteerd door de keten naar O2. Dit
genereert een proton motive force die leidt tot ATP
synthese door ATPase. Zie plaatje.
Nitrificatie zijn twee processen:
- Oxidatie van ammonium (NH3) naar nitriet
(NO2-)
- Oxidatie van nitriet naar nitraat (NO3-)
Deze functioneren als energiebron/elektronen
donor.
NH3 en NO2- worden geoxideerd door het
nitrificeren van bacteriën tijdens nitrificatie. Twee
groepen bacteriën werken samen om ammonia
volledig te oxideren naar nitraat. Sleutel enzymen
2016/2017
zijn:
- Ammonia monooxygenase
- Hydroxylamine oxidoreductase
- Nitriet oxidoreductase
Uit
deze
reactie
komen
alleen
kleine
energieopbrengsten. De groei van nitrificerende
bacteriën is dan ook heel traag.
Ammonia-oxiderende bacteriën oxidiceren NH3 door
ammonia monooxygenase, waarbij NH2OH en H2O
gevormd wordt. Hydroxylamine oxidoreductase
oxidiceerd daarna NH2OH naar NO2-. Elektronen en
protonen worden gebruikt om ATP te genereren. Zie
plaatje.
Nitriet oxiderende bacteriën oxideren nitriet (NO2-)
door het enzym nitriet oxidoreductase naar nitraat
(NO3-). Elektronen en protonen worden gebruikt om
ATP te genereren. Zie plaatje.
Autotrofe organismen gebruiken licht of een
anorganische verbinding als energiebron. De meest
voorkomende manier van CO2 assimilatie is die via de
Calvin-cyclus (met RuBisCo als sleutelenzym).
Anaerobe respiratie is het gebruik van een elektronen
acceptor die niet zuurstof is. Voorbeelden zijn nitraat
(NO3-), ferrisch ijzer (Fe3+), sulfaat (SO4 2-),
carbonaat (CO3 2-) en verschillende organische
stoffen. Hierbij komt echter minder energie vrij dan bij
aerobe respiratie (zie redox toren). Het is afhankelijk
van het elektronen transport, generatie van de proton
motive force en ATPase activiteit.
Lecture 10
Fermentatie en gisting
Bij ademhaling zijn exogene elektronen acceptoren aanwezig voor de oxidatie van elektronen
donoren. Dit zijn meestal zuurstofmoleculen. Bij fermentatie is de donor en het acceptor hetzelfde
molecuul (of het volgt uit elkaar). Als er geen elektronen acceptoren aanwezig zijn kunnen
verbindingen worden gefermenteerd. ATP synthase is dan compleet afhankelijk van fosforylatie. De
redox balans wordt bereikt door fermentatie producten.
Als een organische verbinding wordt opgenomen in een cel en wordt afgebroken door bijvoorbeeld
glycolyse, komt ATP vrij. Door de vorming van NADH van de redox cyclus wordt een fermentatie
product gevormd. Dit kan bijvoorbeeld melkzuur uit pyruvaat zijn. De meeste fermentaties kunnen
alleen maar werken omdat er onderweg intermediaten zijn die energie rijk zijn. Behalve het maken
van melkzuur en ethanol kan er ook NAD+ en H2 gevormd worden als fermentatie product. Hierbij is
dan een ferredoxine (donor) betrokken en een hydrogenase.
Homofermentatieve fermentatie maakt uitsluitend melkzuur.
In herkouwers kan in het rumen cellulose worden verteerd door symbiose. Dit komt door microben.
Deze hydrolyseren cellulose naar vrije glucose dat dan wordt gefermenteerd, waarbij allemaal
vetzuren gemaakt worden en gassen zoals CH4 (methaan) en CO2. De vetzuren zijn belangrijke
energiebronnen en worden opgenomen door de bloedstroom. De micro-organismen maken ook
aminozuren en vitaminen voor het dier. De microben zelf kunnen ook worden opgenomen als bron
voor eiwitten. De meeste microben in het rumen zijn anaeroob (daarom doen ze aan gisting). Er
zitten zo’n 300-400 verschillende soorten in het rumen.
2016/2017
Ook bij de mens zijn veel micro-organismen aanwezig. Deze hebben een heel belangrijk effect op de
vroege ontwikkeling, zoals op het immuunsysteem. Ook is er waarschijnlijk een groot effect op de
hersenen. De grote hoeveelheid van micro-organismen in menselijke moedermelk kan de oorzaak
zijn voor de grote hoeveelheid micro-organismen in onze darmen. Dit is ook zeer afhankelijk van het
soort voedsel. In de maag zitten 10^4/g bacteriën (pH 2). De pH neemt toe tot een pH van 7, hierbij
zijn dan ook de meeste bacteriën (vlakbij de endeldarm).
Ook kan de samenstelling van de bacteriën duiden op obesitas. Bij muizen was te zien dat de muizen
die te dik werden veel meer methaanvormende bacteriën hadden. Bij gewone muizen wordt er
voornamelijk H2 en vluchtige vetten, bij obese muizen wordt er erg veel CH4 aangemaakt.
Symbiose bij insecten
Micro-organismen kunnen uit het milieu worden opgenomen maar kunnen ook door de ouders
worden doorgegeven. Er zijn ook micro-organismen die alleen maar in een symbiose kunnen leven
(kunnen niet vrij leven). Andere symbionten kunnen wel vrij en zelfstandig leven maar hebben een
voordeel en geven een voordeel aan de host. Meestal zijn dit nutritional, bescherming van de
omgeving en bescherming tegen pathogenen (onze huid zit vol met eigen bacteriën, daardoor
kunnen pathogenen zich niet nestelen). Genomen van insecten symbionten zijn veel kleiner dan
genomen van vrijlevende symbionten.
Termieten kunnen cellulose en hemicellulose afbreken en hebben verschillende magen. Hierin
kunnen micro-organismen aanwezig zijn, zoals bijvoorbeeld cellylotische anaeroben. Deze doen aan
het afbreken van cellulose naar glucose naar acetaat of H2, voornamelijk in de hindgut.
Lecture 11 en 12
De koolstof, stikstof en sulfur cyclus
Koolstof cycleert door de grootste reservoirs van de aarde, zoals de atmosfeer, het land, de oceanen,
sedimenten, stenen en biomassa. De grootte van het reservoir en de turnover tijd zijn belangrijke
parameters voor het begrijpen van de cyclus van de elementen.
CO2 in de atmosfeer is het snelste overgeplaatste koolstof reservoir. CO2 wordt gefixeerd door
fotosynthese van landplanten en marine microben. Het gaat weer terug de atmosfeer in door
respiratie en anthropogene activiteiten (microbiële decompositie is de grootste bron voor CO2 die
vrijkomt in de atmosfeer). Koolstof en zuurstof cyclussen zijn gelinkt aan elkaar. Fototrofe
organismen zijn de fundering van de koolstofcyclus. Oxygene fototrofe organismen kunnen worden
verdeeld in twee groepen: planten en micro-organismen. Planten zijn dominant op het land, microorganismen in het water.
Een guild is een populatie microbiëlen met dezelfde metabolische actviteit. Meerdere guilds vormen
microbiële gemeenschappen die op macro-organimsen en abiotische factoren in het ecosysteem in
werken. Een niche is een habitat dat wordt gedeeld door een guild. Het geeft nutriënten en goede
condities voor groei.
Biogeochemie: studie van biologisch gestuurde chemische omzettingen. Een biogeochemische cyclus
bepaalt de transformaties van een sleutel element van biologische of chemische middelen. Dit gaat
meestal via oxidatie-reductie reacties. Microben spelen een kritieke rol in energie transformaties en
biogeochemische processen die resulteren in de recycling van elementen naar levende systemen.
In veel gematigde meren wordt de waterkolom
gestratificeerd in de zomer. Zie plaatje. In rivieren is dit
minder door snel stromend water. Kan nog steeds te weinig
zuurstof hebben door hoge inputs van:
- Organisch matteriaal van het riool.
- Agricultureel en industriële vervuiling
Biochemical oxygen demand (BOD) is de microbiële
zuurstof-gebruikende capaciteit van een hoeveelheid
water.
Fotosynthese en respiratie zijn onderdeel van de redox
cyclus.
2016/2017
Fotosynthese: CO2 + H2O (CH2O) + O2
Respiratie: (CH2O) + O2 CO2 + H2O
De twee grootste eindproducten van decompositie zijn CH4
en CO2. CH4 is een potentieel broeikasgas. Een beetje
methaan gaat in de atmosfeer. Zie plaatje voor de redox
cyclus van koolstof.
Methanogenese staat centraal in koolstof cycling in
anoxische milieu’s. De meeste methanogenen reduceren
CO2 naar CH4 met H2 als elektronen donor; sommigen
kunnen andere substraten naar CH4 reduceren, zoals
acetaat. Methanogenen werken samen met ‘partners’
(syntrofen) die hen nodige substraten geven.
Methanogenische symbionten kunnen worden gevonden in
protisten. Het is mogelijk dat endosymbiotische
methanogenen voordeel hebben van protisten door het
consumeren van H2 gegenereerd van glucose fermentatie.
Biotische processen laten meer CH4 vrij dan abiotische
processen.
N2 is de meest stabiele vorm van stikstof en is een groot
reservoir. Alleen een paar prokaryoten hebben de
mogelijkheid om N2 te gebruiken als een cellulaire
strikstof bron (stikstof fixatie). Denitrificatie is de reductie
van nitraat naar stikstofgas producten en is het grootste
mechanisme waarbij N2 biologisch wordt geproduceerd.
Ammonia dat geproduceerd is door stikstof fixatie of
ammonificatie kan worden geassimileerd in organisch
materiaal of geoxideerd in nitraat.
Zwavel transformaties door micro-organismen zijn
complex. Het grootste deel van zwavel op aarde zit in
sedimenten en stenen als sulfaat en sulfide mineralen,
zoals gypsum en pyriet. De oceanen zijn de meest
significante reservoir van zwavel (als sulfaat) in de
biosfeer.
Hydrogeen sulfide is een belangrijk vluchtig zwavel gas dat
is geproduceerd door bacteriën via sulfaat reductie of
uitgescheiden door geochemische bronnen. Sulfide is giftig
voor veel planten en dieren en reageert met heel veel
metalen. Zwavel oxiderende chemolithotrofen kunnen
sulfide oxideren en elementair zwavel
bij
oxische/anoxische
raakvlakken.
Organische
zwavel
moleculen kunnen ook gemetaboliseerd worden door
micro-organismen. De meest voorkomende organische
zwavel molecuul is dimethyl sulfide (DMS). Dit wordt alleen
geproduceerd in mariene omgevingen als een degradatie
product van dimethylsulfoniopropionate (een alg osmolyt).
DMS kan worden getransformeerd via een aantal
microbiële processen.
2016/2017
Elzenga
Vaatplanten zijn 450 miljoen jaar geleden ontstaan uit algen. Dit heeft tot landleven geleid.
- Nutrientenopname uit de bodem ipv uit het water
- Vorming van wortels
- Hele leven op dezelfde plaats
- Omgeving veel minder constant dan in de oceaan (elementen)
Planten moeten flexibel met hun omgeving om kunnen gaan. Een groot verschil tussen dier en plant
is dat een plant veel meer door het externe milieu wordt beïnvloed dan een dier. Dit uit zich doordat
een plant totipotent is (een cel kan nog tot alles uitgroeien). Een ander verschil is open of
ongedetermineerde groei. De meeste planten kunnen altijd doorgroeien, dieren niet.
Planten die eerst blad hebben en daarna gaan bloeien reageren op de daglengte. Planten
(voornamelijk voorjaarsplanten) die eerst met de bloem omhoog gaan reageren op de temperatuur.
Een éénjarige plant kan je door laten groeien zolang je er voor zorgt dat ze niet bloeien. Een plant die
maar één keer bloeit noemen we monocarpen. Dit zijn meestal éénjarige planten, maar niet altijd.
Meristemen komen altijd uit het cambium, in de top van de spruit en onder aan de wortel. Hierin zit
boven in een germ line: al het genetisch materiaal van de plant waar uiteindelijk de sperma en
eicellen van de plant worden ontwikkeld. Deze delen zo min mogelijk, zodat de genen zo goed
mogelijk blijven.
De voordelen van op het land leven in plaats van in het water:
- Water diffundeert veel langzamer dan lucht (minder beweging in water).
- (vroeger) minder concurrentie.
- Meer zonlicht
- Te veel zuurstof waardoor je fotorespiratie krijgt (zuurstof gaat te langzaam weg).
Nadelen:
- Je staat vast.
- Je bent afhankelijk van je milieu.
- Je staat direct in contact met de elementen (water beter gebuffert).
- Meer moeite voor voedingsstoffen (wortels etc.).
- Zwaartekracht, waardoor je stevigheid nodig hebt.
Apicale meristemen zitten aan de stengel en worteltop. Deze zorgen voor de groei. Ze differentiëren
zich in eerste instantie in drie meristemen:
- protoderm
- procambium (secundaire diktegroei)
- Vulweefsel meristeem (ground tissue)
Verder naar beneden heb je nog een 4e
meristeem:
- Kurk cambium (alleen bij houtachtige
planten)
In de wortel heb je:
- Apicale meristeem
- Wortelmutsje
- 3 secundaire meristemen
Wortelharen zijn uitgroeisels van de epidermis zonder dat er celdeling tussen zit (het is dus maar één
cel). Het is dus wat anders dan zijwortels! Bij secundaire diktegroei (door het laterale
meristeem(zijknop)) heb je een verschil tussen bedekt en naaktzadigen, voornamelijk in het xyleem
(water en mineralen!). Bij bedektzadigen (angiospermen) heb je een aantal grotere buisjes:
houtvaten. Naaktzadigen hebben alleen kleine buisjes (tracheiden). Door deze houtvaten zijn
angiospermen waarschijnlijk succesvoller.
2016/2017
Planten hebben drie typen ground tissues:
- Parenchym (fotosynthese, opslag, etc. veel cytoplasma)
- Collenchym (levende cellen, lijken op sklerenchym, kleinere celwand dan sklerenchym maar
nog steeds heel dik)
- Sklerenchym (zit in de stengels om de vaten, hele stevige cellen, dode cellen, bijna alleen
maar celwand, voor versteviging of bescherming)
De bandjes van Casparie zitten om het xyleem en floëem en dwingt water en nutriënten via het
cytoplasma de plant binnen te komen. Hierdoor heb je actief transport.
Fysiologie van de spruit kennen, zie plaatjes.
Externe invloeden
Het effect van verdamping op de temperatuur van een blad:
De door verdamping van de bladeren veroorzaakte temperatuurverschillen kunnen worden gebruikt
voor een schatting van de gezondheidstoestand van de vegetatie. De huidmondjes reguleren de
gasuitwisseling (O2, CO2 en H2O) tussen het blad en de omgeving. Hierdoor heeft de plant dus
controle over de verdamping. De snelheid van de uitwisseling hangt af van het concentratieverschil
(diffusie). De opening van de huidmondjes wordt gereguleerd door:
- CO2 concentratie
- Licht
- Het hormoon ABA
- Endogene ritme
Veranderingen in de waterpotentiaal van bodem, water en blad zie je over een periode van 24 uur
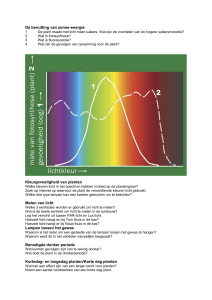
terug in een schommeling. Dit weerspiegelt de mate van verdamping. De formule is:
Mu* + RTln a + VP + zFE + mgh (in J/mol)
R = gasconstante
T = temperatuur in Kelvin
a = activiteit (concentratie * activiteitcoefficient)
V = partieel molal volume
P = druk verschil ten opzichte van 0.1 MPa (=1 atmosfeer)
z = lading
F = Faraday constante
E = electrische potentiaal (V)
m = mol massa
g = zwaartekracht constant
h = hoogte (m)
2016/2017
Als het over de chemische potentiaal gaat ipv water potentiaal valt zFE weg (water heeft geen lading)
en als het over kiemplantjes gaat valt mgh weg (te klein).
Planten die onder erg droge omstandigheden leven zijn beperkt in hun vermogen om door
verdamping de temperatuur laag te houden. Deze hebben zich op andere manieren aangepast:
- Beperken van opname van straling
- Gereduceerd bladoppervlak
- Reflectie van straling (grijze laag-haren, cuticula, etc)
- Aanpassing van de temperatuur-gevoeligheid van de enzymen
Water is ongeladen. Het “reageert” echter wel op
ladingsverschillen (door diffusie), waardoor je verschillende
potentiaal verschillen krijgt. Als een cel heel weinig
voedingsstoffen oid in zich heeft, gaat het water uit de cel
om de osmotische waarde gelijk te krijgen. Dit heet
hypertonisch, de cel is dan geplasmolyseerd. Als de lading
binnen en buiten de cel gelijk is heet dat isotonisch en
noemen we de cel flaccid. Als de lading binnen de cel groter
is dan buiten de cel gaat het water in de cel vacuole en heet het hypotonisch. De cel noemen we dan
turgid (turgor).
De verschillen in waterpotentiaal (of componenten daarvan) zijn de drijvende kracht om alle delen
van de plant van water te voorzien. Van de bodem, via de verschillende compartimenten van een
plant, tot atmosfeer verplaatst het water zich van hogere naar lagere potentiaal. Het watertransport
gaat via het xyleem. Dit gaat voornamelijk van wortel naar scheut. Transport van water en nutriënten
en transport van hormonen (signaal). Transport in het xyleem gaat via de tracheiden in alle
vaatplanten. In angiospermen heb je ook nog houtachtige vaten (meestal dode cellen). De drijvende
kracht voor xyleem transport is:
1. Worteldruk (slechts sterk genoeg om de xyleemvaten weer te ‘vullen’, voor berekening zie
dia 64)
2. Capillaire opstijging
a. Cappilaire opstijging hangt af van de diameter van de buis (een normale afmeting
van xyleemvaten is 20 micrometer). In xyleemvaten zou capillaire werking slechts
kunnen leiden tot een hoogte van 0.75 m. De capillaire werking van de celwanden is
echter veel groter (de afstand tussen de polymeren in een celwand is slechts 5
nanometer).
3. Verdamping
a. Een drukbom wordt gebruikt om de waterpotentiaal in het xyleem van een stengel te
meten. Als gelijktijdig de waterpotentiaal in het blad, de verdampingssnelheid en de
stroom door de stengel wordt gemeten, dan kan worden afgeleid dat de stroom door
het xyleem wordt veroorzaakt door de negatieve waterpotentiaal in het blad.
In een sequoia is de druk veroorzaakt door de hoogte van de waterkolom in de stam. In een boom
van 100 m hoogte is dit dus 1 MPa. Om de weerstand van de vaten te overwinnen en waterstroom
op te wekken is een extra druk nodig van nog eens 0.02 MPa/m, dus 2 MPa bij een boom van 100 m.
een sequoia moet en negatieve waterpotentiaal van -3 Mpa in het bladmesophyl in stand houden.
Het transport van assimilaten van het blad naar de rest van de plant vindt plaats in het floëem. Hierin
is transport van koolhydraten (sucrose), aminozuren en hormonen (signaal). Het gaat van volwassen
bladeren (source) naar groeiende bladeren, wortels, vruchten en opslagorganen (sinks). Het floëem
bestaat uit zeefelementen en begeleidende cellen (companion cellen).
- Zeefelementen zijn levende cellen (maar heel uitzonderlijke cellen: ze bevatten alleen
mitochondria, de plasmamembraan en het endoplasmatisch reticulum).
2016/2017
-
-
Voor het functioneren van de zeefvaten is de associatie van de begeleidende cellen
essentieel.
Transport in de zeefvaten vindt plaats door ‘mass flow’.
De drijvende kracht voor deze ‘mass flow’ wordt opgewekt door het accumuleren van
sucrose in de source organen in de zeefvaten en het water dat het zo opgewekte osmotische
gradient volgt.
In de meeste angiospermen in de gematigde streken is de belading van het floëem
apoplastisch.
Transport eiwitten transporteren de sucrose (en aminozuren) vanuit de apoplast in de
begeleidende cel.
Tranport van cel tot cel:
- Via de plasmodesmata worden kleine moleculen van de ene naar
de andere cel getransporteerd.
o Plasmodesmata zijn kanaaltjes gevormd door de
plasmamembraan waardoor een ‘streng’ ER loopt.
- Transport is via diffusie.
- Plasmodesmata kunnen controle uitoefenen.
De drijvende kracht in de floëemvaten is de drukgradient die wordt opgebouwd door water dat de
suikers die in en uit het floëem worden geladen osmotisch volgt.
Doordat de xyleem- en floëemvaten dicht tegen elkaar aanliggen in de vaatbundels is uitwisseling
van water mogelijk (circulatie).
Oriëntatie in de ruimte
Reacties van planten en dieren zijn verschillend.
Planten:
- Trage reacties (voornamelijk op veranderingen in de omgeving)
- Deze reacties zijn vaak energiezuinig
Dier:
- Snelle reacties (predator-prooi gerelateerd: waar dit ook het geval is in planten, dan ook
snelle reactie, blaasjeskruid)
Planten kunnen wel bewegen. We onderscheiden drie soorten bewegingen:
- Nutatie
o Autonome bewegingen van een plant (niet gerelateerd aan stimuli van buiten). Denk
aan het opkomen van kiemplantjes van een zonnebloem. Nutaties zijn ook de
bewegingen die er voor zorgen dat klimplanten de steun die ze nodig hebben kunnen
vinden.
- Nastische reactie
o Slaapbewegingen, die dienen om verdampend oppervlak te verkleinen en om sterke
afkoeling tijdens de nacht te beperken zijn een voorbeeld van een nastische
beweging. Reageert op een prikkel van buiten af maar niet in dezelfde richting als de
prikkel (vleesetende planten).
- Tropisme
o Fototropisme is de gerichte beweging in de richting van de hoogste lichtintensiteit.
Darwin deed in de 19e eeuw voor het eerst onderzoek naar dit verschijnsel. Went
ontdekte dat een chemische signaalstof in de tip van de stengel
werd geproduceerd (auxine). Als deze aan een zijde van de stengel
wordt aageboden, dan wordt aan die zijde de groei gestimuleerd.
Door licht krijg je dus een assymetrische verdeling van auxine.
Thigmo-morfogenese is als een plant door aanraking vertraagd groeit/bloeit.
2016/2017
In de beschaduwde kant van een plant gaat meer auxine zitten waardoor deze kant harder gaat
groeien, waardoor de plant krom groeit (naar het licht toe). Auxine wordt doorgegeven via polair
transport. De celwand wordt in bepaalde cellen dan minder sterk doordat actieve enzymen
(gestimuleerd door de ATP H+ pomp) verbindingen doorknippen in de celwand. Auxine is een planten
hormoon en grijpt actief in op de plantengroei (op de richting).
Nastische bewegingen: denk aan blaasjeskruid (vangen van vliegen). Dit is een hele snelle beweging.
Nastische bewegingen komen voornamelijk voor bij vleesetende planten, omdat ze niet in een
bepaalde richting reageren. De signaaltransductie van kruidje roer me niet is elektrisch (bijna
dierlijk). Voor signiaaltransductie en reactie zijn de volgende elementen nodig:
- Perceptie (sensor)
- Transductie (versterking/integratie)
- Adaptatie (respons)
Veel bewegingen zijn turgor-afhankelijk, zo ook de beweging van kruidje roer me niet. Andere
bewegingen zijn bijvoorbeeld:
- Stomata
- Slaap bewegingen van bladeren
- ‘solar tracking’ zonnebloemen
Turgorveraderingen worden veroorzaakt door afgifte of opname van vooral kalium en chloride ionen.
Interne regulatie
Planten hormonen spelen een belangrijke rol bij het integreren van verschillende onderdelen van de
plant. Vorm en functie van multicellulaire organismen zijn afhankelijk van effectieve communicatie
tussen cellen, weefsels en organen. In hogere planten hangt de regulatie en coördinatie van
metabolisme, groei en morfogenese af van de uitwisseling van chemische signalen. Dit kunnen
bijvoorbeeld ook suikers zijn (al zijn dat dus geen hormonen). Hormonen zijn:
- Een natuurlijk voorkomende verbinding die wordt gesynthetiseerd in een deel van de plant
die wordt getransporteerd naar een ander orgaan waar het een specifieke werking heeft in
een zeer lage concentrate.
o Speciaal gesynthetiseerd in ander deel van de plant dan waar het heen gaat.
o Natuurlijk voorkomende verbinding
o Zeer lage concentratie
- Ze kunnen groei bevorderend of remmend zijn.
- De werking kan rechtstreeks zijn, ze kunnen eiwitten activeren
- Groei stimuleren:
o Auxine
o Gibberilline
o Cytokinine
- Groei remmen:
o Abscisine zuur
o Ethyleen
- Andere signaal stoffen (bijvoorbeeld waarschuwende stoffen/beschermende stoffen):
o Jasmonzuur
o Salicylzuur
o Systemin
Auxine is bij een heleboel betrokken:
- Tropieën en nastieën
- Celstrekking
- Apicale dominantie
- Stimulering celdeling (verwondingen)
- Weefsel organisatie (organen)
- RNA en eiwit synthese
2016/2017
-
Uitstel van bladval
Interactie met ethyleen (met name bij de bladval)
Voor geordende groei moeten niet al teveel meristemen gelijktijdig actief zijn: apicale dominantie.
Het topje van het apicale meristeem heeft de hoogste concentratie auxine (daar wordt het ook het
meeste gesynthetiseerd). Als je het dominante apicale meristeem er af knipt gaan de zijknoppen ook
groeien. Auxine zorgt er voor dat alle andere knoppen niet uit lopen. De concentratie is echter in het
apicale meristeem het hoogst; daarom zijn verschillende weefsels verschillend gevoelig voor auxine.
Het apicale meristeem is heel ongevoelig voor auxine (daarom groeit die wel), de laterale
meristemen zijn een stuk gevoeliger voor auxine. Auxine is ook betrokken bij de ontwikkeling van
fruit, het wordt geproduceerd in de zaadjes. Auxine zorgt dus vanuit de zaden voor de rijping van de
bloembodem.
Gibberelline is ook belangrijk:
- Celstrekking
- Celdeling
- Enzym inductie
- Bloei-lange dag
- Breekt knop- en zaadrust
Is in Japan ontdekt, doordat sommige rijstplanten heel hoge stengels maakten (en altijd omvielen).
Gibberelline werkt ook op andere planten, zoals op erwten. GA geeft geen ‘acid growth’ zoals auxine.
Komt voor in volwassen weefsel en is niet alleen celstrekking maar ook celdeling. GA zorgt ervoor dat
de spruit op het goede moment uitkomt uit het zaad. Dit komt
vanuit het embryo, wat zich dan verspreid over het zaad. Dit maakt
dan amylase aan waardoor het zetmeel wordt omgezet in suiker,
waardoor het kiemplantje kan gaan groeien/eten.
Cytokinine:
- Celdeling
- Celdifferentiatie
- Breekt knoprust
- Werkt apicale dominantie tegen
- Voorkomt senescentie (veroudering)
- Reguleert eiwitsynthese
Als cytokinine ongeregelt aanwezig is leidt dat tot vorming van woekeringen van ongedifferentieerde
cellen. Dit kan gebeuren door een infectie met Agrobacterium (zorgt voor een verhoogde synthese
van cytokinine). Heksenbezems in berken zijn het resultaat van het ongecontroleerd uitlopen van
okselknoppen. Cytokinine kan ook de veroudering tegen gaan, waardoor je bijvoorbeeld in een
tabaksplant meer goede bladeren behoud (de bladeren in de schaduw blijven ook).
Abscisinezuur (ABA):
- Controle over huidmondjes
- Remming van embryo ontwikkeling
- Houdt knop- en zaadrust in stand
- Abcissie van blad en vrucht
- Staat bekend als (water)stresshormoon
Waarom wil je je bladeren kwijt?
- Geen last van wind
- Compost
- Pathogenen kwijt
Als planten ongevoelig zijn voor Abscisinezuur is de kiemrust afwezig. In een maiskolf zie je dat de
zaden al kiemen aan de maiskolf (niet goed).
2016/2017
Ethyleen:
- Gasvorming (kan daardoor buurplanten beïnvloeden)
- Snelle diffusie
- Interactie met auxine
- Abscissie (samen met ABA) (afvallen van fruit en bladeren)
- Stimulering senescentie (veroudering)
- Fruitrijping
- Nog iets
Ethyleenremmer kan gebruikt worden om bloemen langer te laten bloeien. Antisense ethyleen
synthase remt veroudering van fruit. Dit werden de eerste gemodificeerde producten (tomaten).
Deze konden niet bederven. Bij de auxine-ethyleen interactie zorgen er voor dat een blad geel wordt
en er af valt.