15
Celstofwisseling
2.1Opbouwprocessen in de cel – 16
2.1.1De cel is de kleinste chemische fabriek ter wereld – 16
2.1.2De synthese van een eiwit is als het schrijven van een lang woord, met
aminozuren als letters – 16
2.1.3De eiwitsynthese wordt gestuurd vanuit het DNA in de celkern – 16
2.1.4RNA verbreidt en vertaalt de genetische code – 18
2.1.5De regulatie van de genexpressie – 20
2.1.6Lipiden worden opgebouwd in het gladde ER en verder bewerkt in het
golgi-complex – 21
2.2Afbraakprocessen in de cel – 22
2.2.1De cel splitst stoffen voor (her)gebruik van de componenten of om energie
vrij te maken – 22
2.2.2De splitsing van koolhydraten en vetzuren levert de energie voor alle
celfuncties – 22
2.2.3Enzymen katalyseren biochemische reacties – 24
2.2.4Zuurstof is onmisbaar voor een voldoende vorming van ATP; glucose is
onmisbaar voor een voldoende vetzuurverbranding – 24
L.N. Bouman et al., Leerboek medische fysiologie, Kernboek,
DOI 10.1007/978-90-313-9910-9_2, © 2014 Bohn Stafleu van Loghum, onderdeel van Springer Media BV
2
16
Hoofdstuk 2 • Celstofwisseling
Leerdoelen
2
Na bestudering van dit hoofdstuk en de bijbehorende leerstof op
de website kan men de volgende vragen beantwoorden.
1.Hoe worden op moleculair niveau de erfelijke eigenschappen tot expressie gebracht?
2.Hoe ontstaat RNA? Welke soorten zijn er en wat is van elk de
specifieke functie?
3.Hoe verloopt de eiwitsynthese en hoe wordt deze gereguleerd?
4.Hoe verloopt de citroenzuurcyclus en wat is de ademhalingsketen?
5.Wat zijn lipiden? Waar worden ze gevormd en welke functies
vervullen ze in het lichaam?
Benodigde voorkennis
7 Hoofdstuk 1.
2.1Opbouwprocessen in de cel
2.1.1De cel is de kleinste chemische fabriek ter
wereld
De celstofwisseling omvat vele duizenden chemische reacties die
continu in de cel plaatsvinden. Nieuwe moleculen ontstaan op
verschillende manieren. Dat kan door het ombouwen van uit het
uitwendige milieu opgenomen (= assimilatie) of reeds aanwezige
stoffen, door samenvoeging van kleinere brokstukken tot één
groot molecuul (= anabolisme) of door splitsing van grote moleculen in kleine stukken (= katabolisme). Als het bij dit laatste
proces gaat om afbraak van organische stoffen, bijvoorbeeld de
verbranding van glucose in de mitochondriën, dan spreekt men
van dissimilatie. De chemische basis van deze reacties wordt uitgelegd in intermezzo 2.1
De machinerie die hiervoor nodig is, bestaat uit door de
cel geproduceerde eiwitten die als katalysator fungeren: de enzymen; ze vormen het belangrijkste bestanddeel van het cytoplasma. Veel enzymen komen verspreid in het cytoplasma voor,
maar er zijn ook enzymen die opgehoopt zijn in organellen en
daar dan een specifieke functie bepalen (7 par. 1.1.3). Behalve de
enzymen zijn er nog tal van andere eiwitten met vitale functies.
De eiwitmoleculen in de membraan fungeren in de eerste plaats
als transportmiddel voor uitwisseling van stoffen tussen de extra- en intracellulaire ruimte. Als tweede belangrijke functie
vormen vele membraaneiwitten een aangrijpingpunt (receptor)
voor allerlei van buiten de cel komende stoffen die een reactie
van de cel kunnen oproepen. Intracellulair vormen sommige
eiwitten langgerekte structuren (filamenten) die stevigheid geven aan de cel (cytoskelet) of door samentrekking (contractie)
beweging tot stand kunnen brengen (7 intermezzo 1.4). Een
derde belangrijke functie van eiwitmoleculen is afweer, doordat ze langs chemische weg micro-organismen of tumorcellen
onschadelijk kunnen maken. Ten slotte werken eiwitten ook als
signaalstoffen die rechtstreeks van invloed zijn op naburige cellen (transmitters) of indirect via het bloed worden verspreid
(hormonen).
2.1.2De synthese van een eiwit is als het
schrijven van een lang woord, met
aminozuren als letters
Eiwitten zijn grote moleculen met een molecuulmassa van soms
meer dan een miljoen. Ze bestaan uit een langgerekte keten die,
met soms vele vertakkingen, is opgebouwd uit tussen de 50 en
2000 aminozuurmoleculen. De keten is op een specifieke wijze
tot een kluwen samengevouwen, zodat het eiwit een specifieke
driedimensionale structuur heeft. Deze structuur bepaalt de
werking van het eiwit; verandering van de vorm (conformatie)
van het eiwit leidt tot een verandering of verlies van functie.
Het aantal verschillende aminozuren in de keten is beperkt
tot twintig, zodat zich de vergelijking met een alfabet van twintig letters opdringt. Door deze in verschillende combinaties te
verbinden, zijn talloze verschillende woorden te maken. Dit verklaart hoe het mogelijk is dat de eiwitmoleculen een individu­
specifieke bouw hebben; slechts één letter (aminozuur) verschil
levert soms al een ander eiwit op.
De aminozuren hebben een specifieke chemische structuur
(7 intermezzo 2.2). Ze bestaan uit een korte of lange koolwaterstofketen, waarbij aan het eerste C-atoom (het α-atoom) zowel
een COOH-groep (carboxylgroep) als een NH2-groep (aminogroep) gebonden is (. figuur 2.1A). Het aminozuurmolecuul is
daardoor amfoteer. Dit betekent dat het zowel zure als basische
eigenschappen kan hebben. De zure carboxylgroep kan zich binden aan de basische aminogroep van een ander aminozuur. De
binding die dan ontstaat, noemt men een peptidebinding. Aldus
ontstaat een dipeptide. Naarmate meer aminozuren gekoppeld
worden, vormen zich tri- en tetrapeptiden tot polypeptiden
(. figuur 2.1B). Eiwitten zijn in feite grote polypeptiden. Doordat
eiwitten aan het ene uiteinde een carboxylgroep (van het eerste
aminozuur) hebben en een aminogroep aan het andere uiteinde
(van het laatste aminozuur), zijn ze zelf eveneens amfoteer.
2.1.3De eiwitsynthese wordt gestuurd vanuit
het DNA in de celkern
Net als bij het schrijven de volgorde van de letters bepalend is
voor de vorming van het woord, is de volgorde van de aminozuren in een eiwit bepalend voor zijn werking. Reeds één verkeerd
geplaatst aminozuurmolecuul is voldoende om de functie van
het eiwit teniet te doen. De eiwitsynthese zelf, de aaneenkoppeling van aminozuurmoleculen, vindt plaats in het endoplasmatisch reticulum (ER), maar wordt volledig gedirigeerd vanuit het
chromatinemateriaal dat de door de ouders geleverde genetische
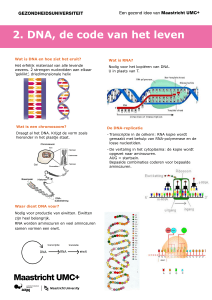
code bevat. Deze code is aanwezig in de structuur van het desoxyribonucleïnezuur (DNA, Eng.: deoxyribonucleic acid).
Nucleïnezuren (DNA) zijn de grootste moleculen in de cel.
Ze zijn opgebouwd uit een lange reeks subeenheden, nucleotiden, die een schijnbaar uniforme opbouw hebben: een stikstofhoudend basenmolecuul, gekoppeld aan een pentose (5C-koolhydraat) dat verbonden is met een fosfaatgroep (. figuur 2.2).
Het essentiële deel van deze eenheid is het basenmolecuul. Er
zijn vier verschillende kernbasen, verdeeld over twee groepen:
A
OH
B
H
carboxylgroep
HO
OH
C
zijketens
R
C
H
polypeptideketen H
O
2
17
2.1 • Opbouwprocessen in de cel
NH2
N
aminogroep
SH
CH3
H3C
CH
CH2
H
H
C
C
H
O
N
C
C
H
O
N
CH2
H
C
C
H
O
CH2
N
C
O
OH
C
H
peptidebinding
N-einde
C-einde
. Figuur 2.1 Bouwstenen van eiwit. A Basale structuur van aminozuren. B Vorming van een polypeptide uit vier aminozuurmoleculen.
Cytosine (C)
A
Thymine (T)
Adenine (A)
C
NH2
NH2
C
N
HN
C
C
N
O
N
O
H
Desoxyribose
C
5
CH2
N
HN
O
1
N
3
H
H
O
OH
4
N
H
Fosfaat
O
OH
C
N
N
H
Guanine (G)
H2N
N
N
CH3
HO
-
O
P
-
O
2
H
B
base
base
O
O
Ri
O
P
O
O
CH2
O
base
H
O
O
P
CH2
O
H
-
O
pentose
O
O
O
P
CH2
O
H
O
fosfaat
O
. Figuur 2.2 A De componenten van DNA. De vier basen C, G, A en T, een suikermolecuul (desoxyribose) en een fosfaatgroep. B Een segment uit een half
DNA-molecuul waarin aan een streng van drie pentosefosfaatdelen drie basenmoleculen gekoppeld zijn.
cytosine (C) en thymine (T), chemisch aangeduid als pyrimidinebasen, en adenine (A) en guanine (G), behorend tot de purinebasen (. figuur 2.2A).
Het DNA bestaat uit twee om elkaar heen gewonden lange
ketens van nucleotiden die via de 3e en 5e positie in de pentosering aan elkaar gekoppeld zijn (. figuur 2.2B). De twee
DNA-ketens zijn via hun kernbasendelen door waterstofbruggen (7 intermezzo 2.1) aan elkaar gekoppeld. Hierdoor ontstaat
het beeld van een wenteltrap (dubbele helix) met de gekoppelde kernbasen als sporten (. figuur 2.3). Per definitie wordt
het einde van de nucleotideketen met de ongebonden 3´-positie in de pentose als 3´-einde (Eng: three prime) aangeduid,
18
Hoofdstuk 2 • Celstofwisseling
2
...
...
A
T
...
...
...
C
G
...
... T
A
suiker-fosfaatskelet
...
T ...
...
C ...
...
thymine (T)
...
T ...
G
...
...
...
G
A
C
A
adenine (A)
cytosine (C)
...
T ...
guanine (G)
G
desoxyribosesuiker
fosfaat
...
...
A
...
...
...
C
...
...
...
G
T
...
...
...
C
A
waterstofbrug
e­ iwitten. Terwijl het humane genoom een gelijke component
van de moeder en de vader bevat (7 H. 23), is het mitochondriale genoom exclusief van maternale origine.
Aan het begin van de celdeling worden de verbindingen tussen de om elkaar gewonden ketens verbroken, zodat de twee
strengen uiteen kunnen wijken (. figuur 2.4A). Langs deze beide
strengen wordt door speciale enzymen, de DNA-polymerasen,
nieuw DNA met complementaire kernbasen gevormd. Als de
bestaande streng bijvoorbeeld een volgorde heeft van TACTGC,
zal de nieuw te vormen streng daar een basenvolgorde van ATGACG vertonen, identiek aan de situatie vóór de splitsing. Dit
geldt op complementaire wijze ook voor de andere helft van het
DNA-molecuul, zodat uiteindelijk na koppeling en herstel van
de ruimtelijke rangschikking in de dubbele winding (helix) nu in
spiegelbeeld twee identieke DNA-moleculen ontstaan (= replicatie), die vervolgens over de twee dochtercellen zullen worden
verdeeld. Dagelijks treden in iedere cel duizenden kleine beschadigingen van het DNA op. Er vindt daarom continu reparatie
van het DNA plaats om te zorgen dat de DNA-code intact blijft
(7 intermezzo 2.3). De volgorde van de basen is de genetische
code, want deze bevat alle informatie die nodig is om de synthese van eiwit en andere celproducten tot in detail te besturen. Wij
beperken ons hier tot de zeer grote lijn van dit primaire levensproces, en staan kort stil bij de manier waarop de eiwitsynthese
door het DNA wordt bestuurd.
2.1.4RNA verbreidt en vertaalt de genetische
code
. Figuur 2.3 Structuur van DNA. Een keten van gefosforyleerde suikermoleculen vormen de gedraaide stijlen van de DNA-ladder, de paren van
complementaire basen A-T en G-C de sporten.
terwijl het DNA-uiteinde met de ongebonden 5´-positie als
5´-einde (Eng: five prime) wordt aangemerkt. De koppeling
tussen nucleotiden is alleen mogelijk tussen adenine en thymine (A-T) en tussen guanine en cytosine (G-C), zodat alleen
deze paren kunnen voorkomen. Die specificiteit, aangeduid
als het complementariteitsprincipe, maakt het mogelijk dat
bij de celdeling het DNA zichzelf exact kopieert. In de celkern is het DNA rond histoneiwitten gewikkeld; deze nucleosoomeenheden liggen nauw gerangschikt en vormen samen
de chromosomen (. figuur 2.4A, B). Het DNA dat zich in de
chromosomen bevindt, bevat specifieke gebieden die de codes
voor eiwitten bevatten, de zogenoemde genen. Het grootste
deel van het DNA (ongeveer 97%) bevat geen genen en wordt
‘niet-coderend’ DNA genoemd. Ook binnen het gen bevinden
zich gebieden die niet coderen. Deze worden introns genoemd;
zij liggen tussen de coderende gebieden die exons worden genoemd (. figuur 2.4C). Het volledige DNA in de celkern van
humane cellen wordt het humane genoom genoemd en bevat bij benadering ongeveer 20.000-22.500 genen. Behalve het
DNA in de celkern wordt er ook nog DNA in de mitochondriën van humane cellen gevonden. Dit DNA wordt ook wel het
mitochondriale genoom genoemd; het is veel kleiner dan het
humane genoom (ongeveer 1% van het totale DNA) en zorgt
voor de productie van een klein deel van de mitochondriale
De selectie van aminozuren en de koppeling van de aminozuren
aan het gewenste eiwit gebeurt in de ribosomen. Ribosomen zijn
complexe clusters van meer dan 50 eiwitten die aan de hand van
de code van het boodschapper-RNA de eiwitsynthese katalyseren. Hieronder beschrijven wij hoe de informatie uit de genetische code van het DNA via RNA (transcriptie = overschrijving)
naar de ribosomen wordt doorgegeven en hoe daar de aminozuurselectie (translatie = vertaling) plaatsvindt.
Voor beide processen maakt de cel gebruik van ribonucleïnezuur (ribonucleic acid, RNA), dat in opbouw lijkt op het DNA. Het
is eveneens opgebouwd uit een suiker, in dit geval echter ribose
in plaats van desoxyribose, en met in plaats van thymine de base
uracil (U), waarin de CH3-groep linksboven in het T-molecuul in
. figuur 2.2 ontbreekt. Ten slotte bestaat het uit slechts één streng,
die vaak veel korter is dan die van de enorme DNA-moleculen.
Het RNA vormt een onmisbare schakel tussen de genetische informatie in het DNA en de vorming van eiwitmoleculen die kunnen worden beschouwd als een complex snoer van aan elkaar
gekoppelde aminozuren (7 par. 2.1.2). Soort en aantal van de ingebouwde aminozuurmoleculen bepalen welke functie het eiwit
kan vervullen. De genetische informatie van het DNA is het recept voor de opbouw van de eiwitten. Het DNA bevat de code die
aangeeft welke aminozuren aan elkaar moeten worden gekoppeld
en in welke volgorde. De aminozuurselectie berust op het feit dat
het DNA opgebouwd gedacht kan worden uit talloze combinaties
van steeds drie aangrenzende kernbasen (= tripletten), die samen
één bepaald aminozuur representeren; de genetische code bestaat
2
19
2.1 • Opbouwprocessen in de cel
B
verloop in tijd
A
DNA
...
...
C ...
histonen in nucleosoom
G
...
A
T ...
...
... G G
C ...
... T
A ...
G
...
...
... C
...
G
C ...
...
kralenketting
...
... T
A
DNA-splitsing
A
T
C
G
nucleosoom
C
G
G
C
G
C
... A
T ...
...
T ...
... T
A ...
...
...
A
...
...
C ...
T
G
... A
T ...
G
...
T ...
oud
A
nieuwvorming
... T
A ...
T
11 nm
T
A
...
...
G
thymine (T)
adenine (A)
G
G
A
identieke chromatide
DNA dubbele helix van
146 nucleotide paren
... A
T ...
...
G ...
... C
...
... C
...
nieuw
histonen in nucleosoom
A
...
...
C ...
nieuw
C
C
C
A
promotorgebied
oud
(model)
regulatoir
element
DNA van een chromatide
cytosine (C)
guanine (G)
5'
gen
primair
promotorgebied
CAAT
exon
exon
TATA
3'
intron
. Figuur 2.4 A Replicatie van DNA. De vermenigvuldiging van de chromosomen begint met het openbreken van de waterstofbruggen tussen de complementaire kernbasen. Door aankoppeling van complementaire kernbasen en verbinding daarvan met desoxyribosefosforzuur ontstaan twee nieuwe,
identieke DNA-ketens. Deze zullen zich over twee dochtercellen verdelen. B Het DNA is rond een cluster van histoneiwitten gewikkeld (nucleosoom). De
nucleosomen liggen op vaste afstanden van elkaar in het DNA; deze structuur wordt ook wel een ‘kralenketting’ genoemd. Een nucleosoom bevat acht
verschillende histoneiwitten; rond dit cluster histoneiwitten ligt ongeveer anderhalve winding DNA met een totale lengte van 146 basenparen. C De organisatie van een eukaryotisch gen. Vóór het gen bevinden zich het primaire promotorgebied en de regulatoire elementen.
dus uit drieletterwoorden die ieder voor zich een symbool zijn
voor één aminozuur (7 intermezzo 2.2). Het RNA vormt een afdruk van een serie van deze tripletten, waardoor het codewoord
voor een eiwit ontstaat. Bij de vorming van het RNA door het
DNA werkt weer het complementariteitsprincipe van de basenparen, met uracil in de rol van thymine. De paren die gevormd
kunnen worden, zijn dus C-G en A-U.
Het RNA komt in vier vormen voor; elke vorm heeft een
afzonderlijke functie in de eiwitsynthese.
1.
Boodschapper-RNA (= messenger RNA, mRNA). Deze RNAketen ontstaat aan het DNA in de kern, op een manier die
sterk lijkt op de eerder beschreven replicatie van DNA. Ook
nu wordt DNA gespleten, maar alleen over het deel dat de
code voor het te synthetiseren eiwit bevat. Aan de gebroken
sporten van de ladder vormt zich nu RNA, met daarin de
tripletten die complementair zijn aan die van het betreffende DNA-fragment. Deze RNA-tripletten noemt men codons,
omdat ze het codesignaal voor de eiwitsynthese in het cytoplasma vormen. De RNA-keten zal evenveel codons bevatten als het eiwit aminozuurmoleculen moet krijgen, vermeerderd met enkele codons voor het starten en stoppen
van de synthese. Omdat er vier basen zijn, die in combinaties van drie worden gebruikt, zijn er 43 = 64 verschillende
codons mogelijk. Er zijn echter slechts twintig verschillende
aminozuren, zodat er voor de meeste aminozuren meer dan
één codon beschikbaar is (7 tabel 2.2.1 in intermezzo 2.2).
Bovendien werkt de combinatie AUG onder bepaalde omstandigheden als startcodon dat het begin van het ‘woord’
aangeeft en zijn er in de tabel verschillende stopcodons te
vinden die het einde van het woord markeren.
20
2
Hoofdstuk 2 • Celstofwisseling
Zoals reeds beschreven, bevatten humane genen zowel
coderende (exons) als niet-coderende (introns) gebieden.
In eerste instantie wordt het volledige gen door RNApolymerase als RNA gekopieerd. Uit dit zogenoemde ‘preRNA’ wordt door selectie van alleen de coderende stukken
het mRNA samengesteld. Er kunnen uit één pre-RNAmolecuul daardoor verschillende combinaties exons en
dus verschillende mRNA-moleculen gehaald worden. Deze
bewerking maakt dat één gen voor verschillende mRNA’s
en dus eiwitten kan coderen. Nadat het mRNA is gevormd,
diffundeert het uit de kern naar het cytoplasma, waar het
zich hecht aan een ribosoom.
2.
Ribosomaal RNA (= rRNA). Dit is een stabiele structurele
vorm van RNA, die verankerd is in de ribosomen. Ribosomen zweven gedeeltelijk vrij in het cytoplasma en zijn gedeeltelijk aan het ER gebonden, zodat het ER daar een ‘ruw’
oppervlak heeft. De vrije ribosomen produceren eiwitten
die in het cytoplasma opgelost aanwezig blijven. In het
ruwe ER worden eiwitten geproduceerd die een onderdeel
van de celstructuur vormen of die als secretieproduct aan
de buitenwereld worden afgegeven. De werkwijze van het
ribosomale eiwitcomplex is in beide gevallen dezelfde en
betreft het binden van mRNA, dat vervolgens in contact
gebracht wordt met het overdracht-RNA (zie hierna) dat de
juiste aminozuren bevat, zodat de translatie van RNA naar
eiwit kan plaatsvinden.
3.
Overdracht-RNA (= transfer RNA, tRNA). Dit RNA wordt,
net als de beide andere vormen van RNA, in de kern gemaakt en aan het cytoplasma afgegeven. Door binding aan
een specifiek aminozuur zorgt het tRNA voor de translatie
(vertaling) van het mRNA naar eiwit. Dit gebeurt doordat
het tRNA een triplet bevat (anticodon) dat complementair
is aan een codon van het mRNA (7 tabel 2.2.1). Kort gezegd
komt de ‘inbouw’ van aminozuren tijdens de eiwitsynthese
tot stand doordat er voor elk aminozuur een specifiek
tRNA bestaat. In het ribosoom koppelt het tRNA via zijn
anticodon op de juiste plaats aan het complementaire codon van het mRNA. Zo komt nu aan het ribosoom een
rijtje verschillende aminozuren liggen, die daarna aan elkaar
gebonden kunnen worden tot een nieuw eiwitmolecuul.
De koppeling van een bepaald aminozuur aan het bijbehorende tRNA komt tot stand via een enzym, een aminozuurtRNA-synthetase, dat reageert met slechts één aminozuur
en het juiste tRNA.
4.
Micro-RNA. Naast de directe rol van de bovengenoemde
RNA-vormen in het translatieproces bestaan er korte
RNA-sequenties (micro-RNA of miRNA genoemd) die
ongeveer 22 nucleotiden lang zijn; miRNA-sequenties
zijn zogenoemde posttranscriptionale regulatoren. Zij
binden aan mRNA en verhinderen hierdoor de eiwitsynthese; binding van miRNA leidt ook tot snelle afbraak
van mRNA. Inmiddels zijn er meer dan 600 verschillende
miRNA-sequenties bekend die in grote mate de eiwitsynthese reguleren.
Het verloop van de translatie van de genetische code in een eiwitstructuur kan nu, sterk vereenvoudigd, als volgt worden sa-
mengevat (. figuur 2.5). Nadat het mRNA in de kern is gevormd,
beweegt het zich door middel van diffusie door de kernmembraan naar het cytoplasma, waar het zich hecht aan het ribosoom. Dit schuift langs het mRNA, waarbij de opeenvolgende
codons in bewerking komen. Deze bewerking bestaat erin dat
per codon een complex van een tRNA-molecuul met het daaraan gehechte aminozuur wordt aangekoppeld. Vervolgens wordt
het tRNA losgemaakt en het aminozuur wordt aan het reeds
aanwezige naburige aminozuurmolecuul vastgemaakt door vorming van een peptidebinding. Het tRNA gaat terug naar het
cytoplasma, op zoek naar een volgend aminozuurmolecuul; het
ribosoom schuift verder langs het mRNA voor de opbouw van
het volgende stuk van het eiwitmolecuul.
De productie van mRNA bepaalt in hoge mate de aard van
de eiwitten waarover de cel zal kunnen beschikken en daarmee
dus de soort cel die zich zal ontwikkelen. Het is dan ook niet
zo dat al het DNA voortdurend bij de vorming van mRNA is
betrokken. In de eerste plaats is slechts een klein deel van het
DNA bij de transcriptie betrokken. In de tweede plaats staat dit
proces onder nauwkeurige controle. Wij spreken daarom van
het mRNA-expressieprofiel: het pakket mRNA-moleculen dat in
een cel van een bepaald type wordt gevormd. Dit expressieprofiel bepaalt welke functies de cel in volwassen toestand zal kunnen vervullen.
2.1.5De regulatie van de genexpressie
Iedere cel in het menselijk lichaam bevat in zijn celkern de volledige genetische informatie. Deze informatie wordt door de
diverse typen lichaamscellen specifiek gebruikt om de eiwitten
die nodig zijn voor de gespecialiseerde celfunctie aan te maken.
De genetische informatie die door gespecialiseerde cellen zoals spiercellen, levercellen en zenuwcellen gebruikt wordt, loopt
sterk uiteen. Terwijl de genexpressie algemeen door de basale
promotorregio (TATA-box) gestart wordt, bevinden zich vóór
het gen regulerende elementen die de celspecifieke genexpressie
bepalen. Over het algemeen hebben humane genen ongeveer vijf
regulatoire elementen die zich in het promotorgebied bevinden
(. figuur 2.4B). Specifieke kleine eiwitten, de transcriptiefactoren,
activeren de regulerende elementen, waardoor de genexpressie
aanvangt. Het zijn de transcriptiefactoren en epigenetische mechanismen (7 intermezzo 2.4) die in grote mate de differentiatie
van celtypen bepalen.
Bij de assemblage van het mRNA-molecuul wordt vaak maar
een deel van de exons van het gen gebruikt. Het verbinden van
de mRNA-sequenties van de diverse exons wordt ‘splicing’ genoemd en hierdoor kunnen van één gen verschillende eiwitten
afgeschreven worden. Uiteraard draagt dit mechanisme bij aan
de diversiteit van de verschillende eiwitten die een cel kan maken.
Doordat, als onderdeel van de specialisatie, cellen hun vermogen om eiwitten te vormen (men zegt: tot expressie te brengen) selectief toepassen, verschilt het pakket aan eiwitten in verschillende cellen sterk. Deze specialisatie komt tot stand door
regeling van de eiwitsynthese op het vlak van de transcriptie, de
vorming van boodschapper-RNA aan het DNA in de kern.
21
2.1 • Opbouwprocessen in de cel
TRANSCRIPTIE
TRANSLATIE
DNA
A
A U
T U
A
T
C
G G
T
T C
A
T A
G
C A
C
G G
A
C
C
A
T G
G
C
A A
G
U
A
T
1 mRNA wordt
gevormd met
DNA als
sjabloon:
(transcriptie)
aminozuur
2 mRNA verlaat de kern
en hecht zich aan de
ribosomen van het ruwe
endoplasmatisch reticulum;
begin van translatie
mRNA
kernporie
kernmembraan
tRNA
aminozuurtRNAsynthetase
polypeptideketen
Met
Gly
Ser
4 doordat het ribosoom
langs de mRNA-keten
schuift, ontstaat een
keten van aminozuren:
het eiwitmolecuul
Phe
5 vrij tRNA komt terug
naar het cytoplasma
om opnieuw als anticodon een aminozuurmolecuul te markeren
mRNA na
translatie
peptideketen
A A
Ile
Ala
3 aminozuren die gebonden zijn aan het
bijpassend tRNA
(anticodon) zoeken
hun plek (codon) in
het ribosoom
G
tRNA met anticodon
U
A
U
ribosoom
C G G
G C C A U
A U U U C
A G U C C
codon
mRNA voor
translatie
. Figuur 2.5 Transcriptie (links) en translatie (rechts): verschillende fasen in de synthese van een eiwitmolecuul.
2.1.6Lipiden worden opgebouwd in het gladde
ER en verder bewerkt in het golgi-complex
Lipiden, vetachtige stoffen die beter in organische oplosmiddelen (bijv. ether) dan in water oplossen (= hydrofoob), vormen
de tweede groep biologisch actieve stoffen die voor een groot
deel in de cel zelf worden gemaakt uit van buitenaf aangevoerde
componenten (7 intermezzo 2.5). Het is een naar structuur minder homogene groep dan de eiwitten, met veel zeer verschillende functies. Om te beginnen zijn lipiden betrokken bij de
vorming van membranen, om de gehele cel of om de organellen. Dit betreft niet alleen stabiele wandstructuren, maar ook
2
22
2
Hoofdstuk 2 • Celstofwisseling
membraanvorming op plaatsen waar een grote synthetische
of transportarbeid plaatsvindt in samenwerking met allerlei
enzym­eiwitten. Daarnaast zijn lipiden belangrijke boodschapperstoffen (bijv. steroïdhormonen) en zijn ze in de vorm van vet
de belangrijkste opslagmogelijkheid van energie voor het lichaam.
De lipiden, in het bijzonder de lipiden die deel gaan uitmaken van een membraan, worden opgebouwd aan het cytosoloppervlak van het gladde ER en daarna naar de binnenkant van
het ER verplaatst en doorgegeven naar het golgi-complex. Na
koppeling aan membraaneiwit wordt het nieuwe membraanmateriaal opgenomen in de celmembraan of in een van de vele
organellen.
Samenvatting
1.Stofwisseling (= metabolisme) is het totaal van processen
voor opbouw (= anabolisme) en afbraak (= katabolisme) van
stoffen. Als de opbouw opname van stoffen uit het uitwendige milieu inhoudt, gaat het om assimilatie; afbraak van
celcomponenten is dissimilatie.
2.De vorming van stoffen vindt plaats via verschillende chemische reacties die door als katalysator werkende eiwitten, de
enzymen, tot stand worden gebracht.
3.De chemische reacties zijn naar functie verdeeld over de
verschillende organellen van de cel:
–– eiwitsynthese in kern en het ruwe deel van het endoplasmatisch reticulum (ER);
–– lipidesynthese in het gladde deel van het ER;
–– afwerking van secretieproducten in het golgi-complex;
–– oxidatieve splitsing van brandstof in de mitochondriën;
–– opname en afgifte van stoffen door de celmembraan.
4.De eiwitsynthese wordt gestuurd vanuit het genetisch materiaal in de kern. Dit bestaat uit zeer grote DNA-moleculen
die, in de volgorde van de basenparen adenine gekoppeld
aan thymine en guanine aan cytosine, de informatie bevatten over de volgorde van aminozuren in de in het ER op te
bouwen eiwitmoleculen.
5.Deze informatie wordt door boodschapper-RNA (mRNA),
dat door transcriptie een chemische afdruk van het DNA
vormt, met uracil in plaats van thymine naar het ER gevoerd.
Het mRNA wordt daartoe aan een ribosoom gebonden.
6.In het ribosoom vindt de translatie plaats van de code in
een aminozuurvolgorde; aminozuren worden herkenbaar
gemaakt door koppeling aan transfer RNA (tRNA).
7.De belangrijkste groep lipiden zijn de glyceriden, opgebouwd uit glycerol en vetzuur. Men onderscheidt verzadigde en onverzadigde vetzuren door het niet of wel aanwezig
zijn van dubbele koolstofbindingen.
2.2Afbraakprocessen in de cel
2.2.1De cel splitst stoffen voor (her)gebruik van
de componenten of om energie vrij te
maken
Tot de dynamiek van de cel behoort dat naast de voortdurende
aanmaak van celbestanddelen er ook continu materiaal wordt
afgebroken. Wij spreken van turnover. De turnovertijd, die aangeeft hoe lang een bepaald product gemiddeld blijft bestaan, varieert van seconden tot jaren, afhankelijk van de functie van het
product. Structuureiwitten hebben een tragere turnover dan enzymeiwitten. De kortste turnovertijd vertonen de signaaleiwitten die na overdracht van een signaal onmiddellijk onwerkzaam
worden gemaakt.
De afbraak van eiwit is de taak van de lysosomen, een heterogene groep vesikels die alle beschikken over ‘zure hydrolasen’,
splitsende enzymen die werkzaam zijn in een zuur milieu. De lysosomen beschikken zelf over de enzymen die lokaal de H+-concentratie kunnen verhogen. Splitsing van lange vetzuurketens
vindt plaats in de peroxisomen, die hun substraat afbreken door
oxidatie. Naast de afbraak van eiwit door lysosomen vindt een
groot deel van de eiwitafbraak plaats door het ubiquitine-proteasoomsysteem. Het proteasoom is een groot eiwitcomplex dat
zorgt voor de afbraak van beschadigde of overbodige eiwitten.
Deze eiwitten worden gemarkeerd met een groot aantal kleine
peptiden die ubiquitines genoemd worden. De ubiquitineketen
die hierdoor ontstaat, bindt aan het proteasoom en zorgt dat
het eiwit opgenomen wordt in het proteasoom. De proteasomen
breken de geübiquinileerde eiwitten af tot peptiden van zeven
of acht aminozuren lang. De aminozuren die vrijkomen bij afbraak van deze peptiden, kunnen opnieuw gebruikt worden in
de eiwitsynthese.
Behalve het hergebruik van onderdelen is een tweede reden van hydrolytische (d.m.v. binding met water) of oxidatieve
(d.m.v. binding met zuurstof) splitsing van stoffen het vrijmaken
van energie door verkleining van de moleculen. Stoffen die deze
eigenschap hebben, noemt men energierijk. In de voeding zijn
het met name de koolhydraten (bijv. zetmeel) en vet die als energierijke substraten worden aangevoerd. Bij een overschot aan
eiwit in de voeding kan dit ook als energiebron worden benut.
De aminozuren worden dan ontdaan van hun stikstofatomen
(desaminering) en de overblijvende C-H-O-verbindingen kunnen worden omgezet in vet of koolhydraat.
2.2.2De splitsing van koolhydraten en vetzuren
levert de energie voor alle celfuncties
De splitsing van koolhydraten (suikers) en vetzuren in water
(H2O) en kooldioxide (CO2) verloopt grotendeels door verbranding (= oxidatieve splitsing) en daarom noemt men deze stoffen de ‘brandstoffen’ van de cel (7 intermezzo 2.6). Dat is maar
gedeeltelijk juist, omdat tot op zekere hoogte ook hydrolytische
splitsing van koolhydraat mogelijk is zonder gebruik van O2. Wij
spreken daarom van aerobe (= oxidatieve) en anaerobe (= nietoxidatieve) stofwisselingsprocessen. Beide omvatten een aantal
chemische reacties. De aerobe processen zijn de belangrijkste.
Niet alleen is de hoeveelheid energie die daardoor vrijgemaakt
kan worden een veelvoud van die door de anaerobe stofwisseling
kan worden geleverd, maar zij kunnen ook doorgaan zolang er
brandstof is, terwijl anaerobe splitsing na korte tijd moet worden gestaakt door een ophoping van afvalstoffen. Daardoor is
de aanwezigheid van zuurstof een absolute voorwaarde voor het
langer voortbestaan van een menselijke cel.
23
2.2 • Afbraakprocessen in de cel
energieke
fosfaatverbinding
+40 kJ/mol
NH2
N
N
P
P
OCH2
P
NH2
opgenomen
energie
P
N
N
N
N
N
P
OCH2
N
N
N
O
O
H2O +
+ P
-40 kJ/mol
HO
OH
ATP
fosfaat
vrijkomende
energie
HO
OH
ADP
. Figuur 2.6 Chemische structuur van ATP (links) en de vorming uit ADP (rechts). Bij de vorming van ATP wordt energie opgenomen, die bij behoefte
aan energie door splitsing weer wordt afgegeven.
ribose
ADP
N+
N
reductie
O
H
ADP
ribose
NH2
NAD+ + H+ + 2e-
O
oxidatie
H
H
NH2
NADH
. Figuur 2.7 Chemische structuur van het NAD+ (links) en de omzetting
in NADH waarbij het een proton (H+) en twee elektronen (e –) opneemt.
De energie die uit deze reeks processen vrijkomt, kan niet
rechtstreeks voor energievragende celfuncties worden gebruikt,
maar wordt eerst opgeslagen als adenosinetrifosfaat (ATP)
(. figuur 2.6). Deze stof kan worden beschouwd als de universele overdrachtsstof voor energie. Elk energievragend proces in
de cel gebruikt de energie uit ATP, waarbij het gesplitst wordt in
adenosinedifosfaat (ADP) en fosfaat; per grammolecuul (= mol)
levert dat 40 kJ op. Een tekort aan ATP luidt het einde van de cellen in, omdat dan essentiële processen zoals membraantransport
tot stilstand komen.
Het meeste ATP wordt gevormd door een reeks van processen in de mitochondriën, die daarvoor in hun binnenste membraan een groot pakket speciale enzymen en transporteiwitten
bevatten. Deze grote groep enzymen vormt de ademhalingsketen
die energie kan halen uit het transport van elektronen door een
reeks van reacties door complexe enzymsystemen. Uiteindelijk
worden deze elektronen met waterstof en zuurstof verenigd tot
een watermolecuul.
De ademhalingsketen kan als leverancier van de elektronen
niet rechtstreeks koolhydraat of vet gebruiken. Deze stoffen
moeten eerst worden gesplitst in kleinere moleculen, waarbij
elektronen worden vrijgemaakt en overgedragen aan enkele specifiek daarvoor in cytosol en mitochondriale matrix aanwezige
waterstofacceptoren, het NAD (nicotinamideadeninedinucleotide, . figuur 2.7) en het daarmee verwante FAD (flavineadeni­
nedinucleotide). Deze stoffen werken oxiderend doordat zij waterstofatomen opnemen tijdens de enzymatische splitsing van
de brandstoffen.
De glucosesplitsing begint al in het cytosol, door een reeks
van reacties die aangeduid worden als glycolyse (7 intermezzo
2.6). Het eindproduct is pyrodruivenzuur (= pyruvaat), maar belangrijker is dat er tijdens de splitsing van één glucosemolecuul
twee moleculen NADH worden gevormd die in de ademhalingsketen kunnen worden verwerkt. Bovendien worden er bruto vier
moleculen ATP uit ADP gevormd; netto is dat maar twee, omdat
in het eerste deel van de glycolyse 2 mol ATP worden verbruikt
om fosforhoudende tussenproducten te maken.
Voor een verdere splitsing moet het pyruvaat naar het inwendige van een mitochondrion worden getransporteerd, waar
zich de enzymen bevinden, voor een cyclisch verlopende reeks
van reacties, de citroenzuurcyclus, naar zijn ontdekker ook wel
de krebscyclus genoemd (7 intermezzo 2.7). Daar worden per
pyruvaatmolecuul drie moleculen NADH gevormd en 1 mol
FADH2. Het pyruvaat wordt daarbij volledig gesplitst in CO2 en
H2O (. figuur 2.9).
Vet kan de celmembraan niet passeren; het moet eerst gesplitst worden in glycerol en vetzuren. Deze laatste passeren
de celmembraan wel en kunnen dan in het mitochondrion als
brandstof dienen. Aldaar aanwezige enzymen brengen een zodanige splitsing van het vetzuurmolecuul tot stand dat ook zij
eveneens in de citroenzuurcyclus verder kunnen worden afgebroken. Deze voorbewerking noemt men de β-oxidatie.
Aminozuren (7 intermezzo 2.2) worden alleen voor energielevering gebruikt als er een overschot is aan aminozuur en een
tekort aan energie. De eerste stap, die alleen in de lever kan worden uitgevoerd, is het aminozuur ontdoen van de aminogroep.
Daarvoor zijn twee mogelijkheden: transaminering en desaminering. In het eerste geval wordt de aminogroep behouden door
overzetting naar α-ketoglutaarzuur (uit de citroenzuurcyclus),
zodat het aminozuur glutaminezuur wordt gevormd. In het
tweede geval gaan de aminogroepen verloren; ze worden omgebouwd tot ureum en in de urine uitgescheiden. Wat overblijft
wordt omgezet in glucose (gluconeogenese) en kan als substraat
voor de glycolyse dienen.
2
24
Hoofdstuk 2 • Celstofwisseling
2.2.3Enzymen katalyseren biochemische
reacties
2
Zoals reeds eerder aangegeven, zijn enzymen eiwitten die als
katalysator fungeren om biochemische reacties bij normale
temperatuur en druk efficiënt te laten verlopen. Voor zijn functie moet het enzym in contact komen met de beginproducten
van de biochemische reactie (het substraat). Het substraat bindt
dus aan het enzym en vormt zo een enzym-substraatcomplex,
dat aan het eind van de reactie splitst in het reactieproduct en
het enzym. De reactie tussen enzym en substraat wordt weergegeven als:
Ssubstraat + E enzym → ESenzym − substraatcomplex → Pproduct + E enzym
Een bekend voorbeeld van de bovenstaande reactie is de fosforylering van eiwitten. Hierbij wordt een eiwit (het substraat) door
een proteïnekinase (het enzym) van een fosfaatgroep voorzien,
zodat er een gefosforyleerd eiwit (het product) ontstaat. Aan het
eind van de reactie zal het enzym de reactie met de overgebleven
substraatmoleculen herhalen, zodat de omzetting van substraat
in product efficiënt verloopt en het enzym deze omzetting effectief versnelt. Het deel van het enzym waaraan het substraat
bindt, wordt de actieve regio van het enzym genoemd en bepaalt welk substraat aan het enzym kan binden (enzymspecifiteit). Iedere cel bevat ongeveer 4000 verschillende enzymen die
verschillende biochemische reacties katalyseren. De naam van
een enzym wordt algemeen samengesteld uit de naam van het
substraat, aangevuld met -ase; bijvoorbeeld enzymen die vetten
(lipiden) afbreken, worden lipasen genoemd.
2.2.4Zuurstof is onmisbaar voor een voldoende
vorming van ATP; glucose is onmisbaar
voor een voldoende vetzuurverbranding
Alle hierboven genoemde mechanismen om brandstofmoleculen tot CO2 en water te verkleinen en energie daaraan te onttrekken voor de vorming van ATP bevatten één of meer oxidatiestappen die tot stilstand komen als er niet voldoende O2 in de cel
aanwezig is. Alleen de glycolyse biedt een zijweg om kortdurend
anaeroob energie vrij te maken. Het pyruvaat dat aan het einde
van de reactiereeks ontstaat, kan indien nodig waterstofatomen
terugnemen van het eerder gevormde NADH, waardoor melkzuur (= lactaat) ontstaat dat zich ophoopt en niet verder kan
worden verwerkt. Dat is dan het noodzakelijke einde van de
anaerobe stofwisseling die per glucosemolecuul dan netto 2 mol
ATP heeft opgeleverd.
Dat is heel anders als er wel voldoende zuurstof is. Dit komt
niet alleen doordat het pyruvaat nu kan overgaan in de citroenzuurcylus, want die levert per pyruvaatmolecuul niet meer dan
1 mol ATP op, via tussenschakeling van GTP, een verwante energierijke fosfaatverbinding (. figuur 2.9). Maar nu kunnen via de
β-oxidatie vetzuren kunnen worden geoxideerd, wat gemiddeld
per mol vetzuur netto 7 mol ATP oplevert. Dit is op zich veel
lucratiever, doordat nu NADH en FADH2 in de ademhalingsketen kunnen worden geproduceerd. Dit levert per mol glucose
ongeveer 34 mol ATP op en per mol vetzuur gemiddeld 128 mol
ATP. Omgerekend per gram brandstof is de energieopbrengst
per 100 g glucose 20 mol ATP en per 100 g vetzuur gemiddeld
50 mol ATP. Uit dit verschil in energie-inhoud van deze verschillende brandstoffen is te begrijpen waarom het lichaam vet
voor langeretermijnenergieopslag gebruikt en geen koolhydraat.
Glucoseverbranding is echter om twee redenen onmisbaar. In de
eerste plaats om een snel startende arbeid zoals een sprint mogelijk te maken. De voor de aerobe energievrijmaking noodzakelijke verhoging van de zuurstofaanvoer komt daarvoor te traag
op gang, zodat de start is aangewezen op de anaerobe stofwisseling; glucose is het enige substraat daarvoor. De tweede reden is
dat bij een onvoldoende glucoseafbraak er een verstoring van de
citroenzuurcyclus optreedt die ook de vetzuursplitsing verstoort
(7 intermezzo 2.8).
De glycolyse
Glucose is de belangrijkste brandstof voor de synthese van ATP.
De suiker wordt door het bloed aangevoerd, in de cel actief opgenomen en dan met behulp van de energie uit twee ATP-moleculen gebonden aan fosforzuur tot fructose-1,6-difosfaat. Dit
zijn de eerste stappen van een reeks van negen reacties die in het
cytosol optreden, waarin met behulp van evenveel enzymen het
glucosemolecuul in twee stukken breekt, met als eindproducten twee moleculen pyrodruivenzuur (pyruvaat, . figuur 2.8)
en vier moleculen ATP. Dit betekent een nettowinst van twee
moleculen ATP, omdat er twee zijn geïnvesteerd aan het begin
van de reeks.
Ongeveer halverwege de procesreeks vindt een oxidatiestap
plaats waarbij twee H-atomen worden onttrokken en gebonden
aan nicotinamideadeninedinucleotide (NAD), zodat NADH + H+
ontstaat. Als er voldoende zuurstof wordt aangevoerd, kan NAD
deze H-atomen aan zuurstof overdragen waardoor water ontstaat (7 intermezo 2.7). Het NAD blijft dan als waterstofacceptor
beschikbaar. Als er echter onvoldoende zuurstof aanwezig is (anaerobe omstandigheden), draagt het NADH + H+ zijn waterstof
over aan het zojuist gevormde pyrodruivenzuur, waardoor melkzuur ontstaat. Dit kan in de cel niet verder worden verwerkt,
zodat het zich zal ophopen. Hoewel door dit mechanisme dus
ATP gevormd kan worden zonder voldoende zuurstoftoevoer
(anaerobe arbeid), is dat slechts gedurende korte tijd mogelijk,
omdat door de ophoping van melkzuur de pH in de cel sterk
daalt en de eiwitten daardoor hun functie verliezen (acidose).
De citroenzuurcyclus
Als de toevoer van zuurstof voldoende is (aerobe omstandigheid), wordt pyrodruivenzuur aan het einde van de glycolyse
niet gereduceerd tot melkzuur en kan het verder worden afgebroken, waarbij weer een aantal oxidatieve reacties optreedt.
Hiervoor moet het pyrodruivenzuur vanuit het cytoplasma naar
een mitochondrion diffunderen, omdat alleen daar de benodigde enzymen aanwezig zijn. De processen in de mitochondriën
zijn zeer complex en niet in een paar woorden uit te leggen.
Voor een gedetailleerd beeld wordt de lezer verwezen naar een
leerboek biochemie. Hier noemen wij slechts de hoofdpunten.
De mitochondriale splitsing van pyrodruivenzuur uit de glycolyse speelt zich af in twee reactieketens. De eerste heeft een
http://www.springer.com/978-90-313-9909-3