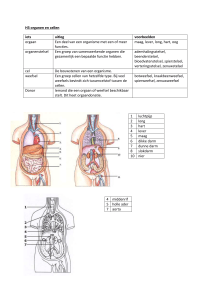
1
Cellen en weefsels
Binnen de humane biologie worden de volgende twee wetenschappelijke disciplines onderscheiden:
– fysiologie: leer der verrichtingen.
Deze wetenschap bestudeert de verrichtingen van het menselijk
lichaam en de functies van de verschillende onderdelen. Dankzij de
recente ontwikkelingen op het gebied van microbiologie en biochemie ontwikkelt de fysiologie zich snel met als gevolg het ontstaan van steeds weer nieuwe onderzoeks- en behandelingsmethoden;
– anatomie: ontleedkunde.
Deze wetenschap houdt zich bezig met het bestuderen van de bouw
van het menselijk lichaam. Zij beschrijft hoe organen ten opzichte
van elkaar zijn gelegen, hoe de bloedvoorziening van deze organen
verloopt etc. Veel anatomische kennis is verkregen door het bestuderen van stoffelijke overschotten. Deze kennis vormt de basis voor
onder andere het lichamelijk onderzoek.
Aangezien functie en bouw van het lichaam nauw met elkaar samenhangen zijn ook de fysiologie en anatomie sterk met elkaar verbonden. Als eerste worden in dit hoofdstuk de kenmerken van het menselijk lichaam als organisme beschreven. Daarbij speelt de homeostase een belangrijke rol. Vervolgens wordt ingegaan op de kleinste
levende eenheden van het menselijk lichaam: de cellen. Cellen met
dezelfde functie vormen weefsels. Ten slotte komt in dit hoofdstuk de
algemene topografie aan bod.
1.1
Kenmerken van het leven
De kenmerken van de levende stof zijn niet gebonden aan de grootte
van een organisme. Bij de elementaire functies van de levende organismen, en dus ook van de mens, worden de volgende kenmerken
onderscheiden:
– metabolisme (stofwisseling); dit is het vermogen om bepaalde stoffen
om te zetten in andere stoffen met als doel het verrichten van een
of andere vorm van arbeid (fysieke arbeid in spieren, elektrische
arbeid in hersenen, chemische arbeid in groei etc.). Ook de functies van het opnemen van stoffen zoals voedsel en zuurstof en het
uitscheiden van afbraakproducten zijn kenmerken van het leven; ze
zijn nodig om de stofwisselingsprocessen in het lichaam mogelijk
te maken.
Het metabolisme bestaat uit twee afzonderlijke deelprocessen: ka-
20
Fysiologie en anatomie niveau 5
–
–
–
–
–
–
tabolisme en anabolisme. Onder katabolisme wordt verstaan het uiteenvallen van grote moleculen in kleinere moleculen. Hiervan is
bijvoorbeeld sprake bij de spijsvertering in het maag-darmkanaal,
maar ook tijdens de verbrandingsprocessen in de cellen. Onder
anabolisme wordt het totaal van alle opbouwreacties verstaan. Als
grondstoffen voor het anabolisme worden tussenproducten van het
katabolisme gebruikt. Zo worden de spiereiwitten bijvoorbeeld
gemaakt uitgaande van aminozuren, die bij de vertering van het
voedsel ontstaan;
groei; hierbij treedt een volumevergroting van het lichaam op
doordat in het lichaam zelf nieuwe bouwstenen worden gevormd.
Groei vindt plaats nadat met behulp van bepaalde stofwisselingsprocessen de hiertoe benodigde stoffen, zoals voedsel en zuurstof,
vanuit de omgeving zijn opgenomen;
voortplanting; hierbij neemt het aantal individuen toe, wat noodzakelijk is voor het voortbestaan van de soort;
adaptatie; dit is het vermogen om zich aan te passen aan veranderde
levensomstandigheden, bijvoorbeeld de toename van het aantal
erytrocyten (rode bloedcellen) bij een verblijf in een omgeving met
een relatief lage zuurstofspanning (hooggebergte);
prikkelbaarheid; dit is het vermogen om te reageren op prikkels
vanuit de buitenwereld of vanuit het menselijk lichaam zelf;
prikkelverwerking; de inwendige reacties blijven vaak niet beperkt tot
een bepaalde plaats maar kunnen door het gehele lichaam worden
doorgegeven en verwerkt;
beweging; het lichaam en de lichaamsonderdelen veranderen van
vorm en plaats.
Bij al deze functies is er ook nog een onderscheid in animale en
vegetatieve functies. Animale functies zijn functies die het lichaam in
staat stellen te reageren op plotselinge veranderingen van de omgeving zoals prikkelbaarheid, prikkelverwerking en beweging. Vegetatieve functies zijn functies die de groei, de ontwikkeling en het voortbestaan van het individu en de soort mogelijk maken. Hiertoe behoren: opname van voedsel en zuurstof, celstofwisseling, uitscheiding,
groei en voortplanting.
1.2
Hiërarchische opbouw
De cel is de kleinste levende zelfstandige eenheid waaruit het organisme is opgebouwd. Het is de fundamentele bouwsteen van het
menselijk lichaam.
Bij meercellige organismen, zoals de mens, is er sprake van een
sterke specialisatie van de cellen, dat wil zeggen dat de aanvankelijk
uniforme cellen zich geleidelijk op verschillende wijzen hebben ontwikkeld met het oog op een bepaalde functie, waartoe ze tevens een
bepaalde vorm hebben gekregen.
Een groep cellen met dezelfde vorm en functie heet een weefsel, bijvoorbeeld spierweefsel en zenuwweefsel. Meestal bevat een weefsel
een karakteristieke tussencelstof.
1
Cellen en weefsels
Een orgaan is opgebouwd uit verschillende, samenwerkende weefsels,
waardoor het geheel een bepaalde functie uitoefent. Het hart zorgt
bijvoorbeeld voor de circulatie van het bloed.
Een orgaanstelsel bestaat uit een groep samenwerkende organen belast
met het uitvoeren van een bepaalde functie; het spijsverteringsstelsel
bijvoorbeeld wordt gevormd door mond, oesofagus (slokdarm),
maag, darmen, pancrea en galblaas.
Het menselijk lichaam, dat uit verschillende samenwerkende orgaanstelsels is opgebouwd, wordt een organisme genoemd.
1.3
Het menselijk lichaam als eenheid
Als gevolg van specialisatie van de cellen van het menselijk lichaam
kan niet iedere cel alle functies uitoefenen; hij is van andere delen van
het organisme afhankelijk.
Voor deze onderlinge samenhang en coördinatie van functies van
verschillende organen en orgaanstelsels zijn transport en regulatie
nodig. Zuurstof en voedsel worden vanuit de omgeving in het lichaam
opgenomen en via het bloedvatstelsel naar de cellen getransporteerd.
Vanuit de cellen worden afbraakproducten óf via het lymfevatstelsel óf
rechtstreeks door het bloedvatstelsel naar de uitscheidingsorganen
vervoerd. Deze zorgen er op hun beurt voor dat de afbraakproducten
uit het lichaam worden verwijderd.
Het reguleren geschiedt door het zenuwstelsel en het hormoonstelsel.
Het zenuwstelsel en het hormoonstelsel zorgen voor de integratie van
de verrichtingen van de organen en de orgaanstelsels; deze stelsels
laten alle organen en orgaanstelsels op de juiste wijze samenwerken.
Ook het op een doeltreffende en snelle wijze reageren op veranderingen in de buitenwereld wordt door deze stelsels geregeld. Het
zenuwstelsel is in staat om signalen met hoge snelheid door het gehele
lichaam te sturen waardoor de werking van de organen (bijv. hart en
longen) plotseling versneld of vertraagd wordt. Het hormoonstelsel
werkt langzamer, wat verklaard kan worden door het feit dat de
hormonen, die door speciale hormoonklieren of bepaalde weefsels
worden afgescheiden, via het bloed elders in het lichaam hun regulerende werking uitoefenen. In veel gevallen hebben de hormonen
een stimulerende werking op bepaalde organen.
1.3.1
homeostase
Met homeostase (homeostase = gelijk blijven) wordt aangeduid het
streven van het lichaam naar het constant houden van het inwendige
milieu van het lichaam, waartoe ook het bloed behoort. Alle systemen
in het lichaam (ademhaling, spijsvertering, uitscheiding etc.) veroorzaken veranderingen in dit inwendige milieu. Doordat deze systemen
ook elkaar beı̈nvloeden worden deze veranderingen meestal snel weer
gecorrigeerd. Het zenuwstelsel en het hormoonstelsel spelen hierbij
een belangrijke rol. Twee voorbeelden maken het principe van homeostase duidelijk.
Iemand die gaat sporten produceert meer koolstofdioxide (CO2) door
de toegenomen verbranding. Vrijwel onmiddellijk neemt dan de
21
Fysiologie en anatomie niveau 5
22
stroomsnelheid van het bloed toe (het hart gaat sneller kloppen) om
dit CO2 naar de longen af te voeren. Ook wordt de ademhaling gestimuleerd zodat het CO2 daarna kan worden uitgeademd. Door dit
compensatiemechanisme zal de CO2-concentratie in het bloed maar
beperkt stijgen.
Na een maaltijd wordt er veel glucose in het bloed opgenomen. Vrijwel direct reageert het lichaam op deze verandering met de productie
van insuline. Dit hormoon stimuleert onder andere de opname van
glucose in cellen, waardoor de concentratie in het bloed weer normaliseert.
In dit boek zal op een groot aantal plaatsen aandacht worden geschonken aan processen, die van belang zijn voor de homeostase. In
dit hoofdstuk worden in dit verband de vochtbalans, de elektrolytenbalans en de regulatie van de pH besproken.
Vochtbalans
De hoeveelheid water die dagelijks bij volwassenen moet worden
opgenomen bedraagt gemiddeld 2,1 liter. Dit gebeurt via dranken en
vast voedsel. Daarnaast wordt er per etmaal nog ongeveer 0,4 l oxidatiewater geproduceerd tijdens het verbrandingsproces in de cellen.
De opgenomen hoeveelheid moet gelijk zijn aan de hoeveelheid vocht
die dagelijks met de urine, zweet, feces en via uitademing het lichaam
verlaat. Er is sprake van een zogenaamde waterbalans of vochtbalans.
Via de huid verdampt 300-400 ml water ongemerkt door uitwaseming. Het totale waterverlies is afhankelijk van de omgevingstemperatuur. Bij warm weer of zware arbeid verliest het lichaam door zweet
meer vocht (tabel 1.1).
Tabel 1.1
Vochtbalans.
vochtverlies
normale temperatuur
warm weer
langdurige zware arbeid
huid
350 ml
350 ml
350 ml
luchtwegen
350 ml
250 ml
650 ml
feces
100 ml
100 ml
100 ml
zweet
200 ml
1 500 ml
5 000 ml
urine
1 500 ml
1 300 ml
600 ml
totaal
2 500 ml
3 500 ml
6 700 ml
Het lichaam van een jongvolwassene bevat 60% water en 40% vaste
stof. Een persoon met een gewicht van 70 kg heeft dus ongeveer 42
liter water. Bij een vochtbalans van 2,5 liter per etmaal zet een volwassene dus ongeveer 6% (2,5/42) van de totale hoeveelheid water
om. Anders gezegd: in ruim zestien dagen is al het lichaamswater
‘ververst’.
Van de 42 liter water in het lichaam van een volwassene is ongeveer
70% intracellulair (30 liter) en de overige 12 liter bevindt zich extracellulair: 3 liter bloedplasma en 9 liter weefselvocht (interstitieel
vocht).
1
Cellen en weefsels
Het percentage water in het lichaam varieert sterk met de leeftijd: een
zuigeling bevat ongeveer 75% water, een bejaarde 45 tot 50%. Hiermee moet bij de dosering van geneesmiddelen rekening worden gehouden. Ook de wateromzet varieert per leeftijdscategorie: een baby
van 6 maanden oud heeft een gewicht van 7 kg. De totale hoeveelheid
lichaamswater is dan ongeveer 4,6 liter. Op deze leeftijd is de vochtbalans ongeveer 1 liter per etmaal, ofwel 22% van het lichaamswater.
Anders gezegd: deze baby ververst in vijf dagen de totale hoeveelheid
lichaamswater: driemaal zo snel als een volwassene. Het risico op
uitdroging is daarom bij jonge kinderen veel groter dan bij volwassenen.
De vochtbalans is sterk gekoppeld aan de elektrolytenbalans (zie
volgende paragraaf ). Water speelt in het lichaam een veelzijdige rol:
bouwstof (het cytoplasma bestaat voor 75% uit water), oplosmiddel
en transportmiddel. Water speelt bovendien een belangrijke rol bij de
warmteregulatie door transpiratie en is dan ook te beschouwen als de
koelvloeistof bij uitstek. Omgekeerd vervoert het water de warmte van
de plaats waar het wordt geproduceerd (lever, spieren) naar de
plaatsen waar warmte nodig is om het lichaam op temperatuur te
houden.
Om een te hoog aantal afvalstoffen in het bloed (‘uremie’, zie intermezzo 1.1) te voorkomen moet een volwassene ten minste 400 ml
urine per etmaal produceren. In de praktijk wordt een veilige ondergrens van 1 000 ml voor een volwassene aangehouden. Bij de regeling
van de vochtbalans spelen enkele hormonen een belangrijke rol. Deze
zullen in andere hoofdstukken uitgewerkt worden.
Intermezzo 1.1 Verstoorde vochtbalans
Wanneer de vochtbalans is verstoord heeft dit ernstige gevolgen
voor de lichaamsfuncties. Een vochtverlies van 10% is al zeer
ernstig; wanneer het vochtverlies meer dan 20% bedraagt, kan
dit dodelijk zijn. Bij sterk vochtverlies zullen de nieren minder
urine produceren om het evenwicht zoveel mogelijk te kunnen
handhaven. Bij extreem vochtverlies (bijvoorbeeld door een
massale bloeding) kan de productie van urine zelfs geheel stoppen. Dit wordt anurie genoemd. Het gevolg is dat de concentratie van allerlei afvalstoffen in het bloed stijgt met ernstige
consequenties (‘uremie’).
Een te groot watergehalte in het lichaam is eveneens schadelijk.
Dit kan bijvoorbeeld het gevolg zijn van het (meestal onder
dwang) in korte tijd drinken van grote hoeveelheden water (hyperhydratie). Waterintoxicatie kan ook het gevolg zijn van een
overmatige afgifte van het hormoon ADH (zie hoofdstuk 7).
Hierdoor ontstaat waterretentie en als gevolg daarvan hyponatriëmie. In gevorderde gevallen kan dit leiden tot verwardheid,
convulsies en eventueel coma.
23
24
Fysiologie en anatomie niveau 5
Elektrolytenbalans
De mens moet dagelijks een bepaalde hoeveelheid mineralen (zouten)
opnemen. In de fysiologie wordt liever de term elektrolyten gebruikt
omdat de zouten in oplossing altijd in ionen (geladen deeltjes) zijn
gesplitst. Er zijn positieve ionen (kationen), zoals H+, Na+, K+ en
Ca2+, en negatief geladen ionen (anionen), zoals Cl- (chloride) en
HCO3- (bicarbonaat).
De nieren (zie hoofdstuk 5) hebben een belangrijke invloed op de
elektrolytenbalans.
Bepaalde elektrolyten heeft het lichaam slechts in beperkte hoeveelheden nodig. Deze worden sporenelementen of micro-elementen
genoemd. Voorbeelden zijn fluor, jood, koper, zink en kobalt.
De functie van de elektrolyten kan in het kort als volgt worden samengevat:
– bouwstof, bijvoorbeeld calcium- en magnesiumzouten in de botten
en ijzer als bouwsteen van hemoglobine (Hb) in de erytrocyten (zie
par. 2.1.5);
– osmolariteit; dit begrip slaat op het evenwicht dat moet bestaan in de
samenstelling van lichaamsvloeistoffen (bloed, lymfe, weefselvocht) en cytoplasma (zie par. 1.6.2);
– bestanddelen van hormonen en enzymen; voorbeelden hiervan zijn jood
in de schildklierhormonen T3 en T4, koper, kobalt en ijzer die
betrokken zijn bij talrijke enzymfuncties;
– impulsgeleiding;
– spiercontractie.
In hoofdstuk 10 is de functie van de elektrolyten meer uitgebreid
beschreven.
pH
Van alle deeltjes die in het bloed voorkomen is het waterstofion (H+)
het kleinste en het meest agressief. Waterstofionen kunnen met
praktisch alle organische verbindingen reageren, waarbij veel schade
kan worden aangericht. Het is daarom van het grootste belang dat de
concentratie waterstofionen in het bloed constant wordt gehouden.
In de scheikunde wordt onder een zuur verstaan: een stof die waterstofionen kan afstaan. Bekende voorbeelden van zuren zijn: zoutzuur,
zwavelzuur, azijnzuur en fosforzuur. De chemische formule van
zoutzuur is HCl, een verbinding van waterstof en chloor. Wanneer
zoutzuur in water wordt opgelost zullen alle moleculen splitsen in
waterstofionen en chloride-ionen. Een dergelijk zuur wordt een sterk
zuur genoemd. De splitsing van zoutzuur kan als chemische reactie als
volgt worden weergegeven:
HCl ? H+ + ClAzijnzuur (hier weergegeven als HAc) is een voorbeeld van een zwak
zuur. Dat betekent dat niet alle azijnzuurmoleculen splitsen. Een deel
van moleculen blijft als HAc in de oplossing. Dit wordt als volgt
weergegeven:
HAc $ H+ + Ac-
1
Cellen en weefsels
Wanneer oplossingen van zoutzuur en azijnzuur van gelijke sterkte
met elkaar worden vergeleken, zullen er dus in de zoutzuuroplossing
meer H+-ionen voorkomen dan in de azijnzuuroplossing.
In zuiver water komen vrijwel geen gesplitste moleculen en dus ook
bijna geen H+-ionen voor. Zuiver water is dus te beschouwen als een
zeer zwak zuur.
Helaas is aan de formule van een stof niet af te lezen of een stof een
sterk zuur is of niet. Zo heeft glucose de formule C6H12O6. Toch is
glucose in het geheel niet zuur, omdat de waterstofatomen onlosmakelijk in het molecuul gebonden zijn.
Een base is een stof die H+-ionen kan binden. Een voorbeeld van een
base is een oplossing van natriumhydroxide (natronloog). De hydroxide-ionen binden waterstofionen volgens de reactie:
OH- + H+ ? H2O
Hydroxide is een sterke base, dat wil zeggen: vrijwel alle aanwezige
waterstofionen worden gebonden. Een voorbeeld van een zwakke base
is fosfaat. De reactie van fosfaat (PO43-) met waterstofionen wordt dan
geschreven als:
PO43- + H+ $ HPO42Een oplossing is zuur als in die oplossing meer H+-ionen voorkomen
dan in zuiver water. Dit kan alleen maar als er een zure stof in het
water is opgelost die H+-ionen heeft afgestaan.
Een oplossing is basisch of alkalisch als er minder H+-ionen in
voorkomen dan in zuiver water. Dit kan alleen maar als er in het water
een basische stof is opgelost, immers die stof heeft H+-ionen uit het
water gebonden.
Om aan te geven of een oplossing zuur dan wel basisch is, is de
grootheid pH ingevoerd. De pH van zuiver water is precies gelijk aan
7,0. Een zure oplossing heeft een pH die kleiner is, een basische
oplossing heeft een pH die groter is dan 7,0. Hoe lager de pH, hoe
zuurder de oplossing (= hoe meer waterstofionen). Een oplossing van
zoutzuur heeft dus een lagere pH dan een vergelijkbare oplossing van
azijnzuur. De pH-schaal is een logaritmische schaal. Dat betekent dat
in een oplossing met een pH van 6,0 tienmaal zoveel waterstofionen
voorkomen dan in een oplossing met een pH van 7,0.
De pH van arterieel bloed (dus in de slagaders) ligt altijd tussen 7,35
en 7,45. Arterieel bloed is dus licht alkalisch. Bij een pH lager dan
7,35 zijn er zoveel meer H+-ionen in het bloed dat eiwitten beschadigd kunnen raken. Een dergelijke situatie wordt een acidose genoemd.
Bij een pH groter dan 7,45 (alkalose) worden er juist H+-ionen van de
bloedeiwitten afgehaald. Ook dit is al heel snel levensbedreigend.
Buiten de arteriële bloedbaan kan de pH wel buiten de grenzen van
7,35 en 7,45 komen. Een bekend voorbeeld is de pH in de maag. Door
de aanwezigheid van zoutzuur bedraagt deze ongeveer 2,0.
25
26
Fysiologie en anatomie niveau 5
Bij de verbrandingsprocessen in het lichaam ontstaan voortdurend
zuren. De belangrijkste zijn koolzuur en melkzuur. De volgende factoren voorkomen dat de pH van het bloed, ondanks de vorming van
deze zuren, te veel gaat veranderen:
– pH-buffers; dit zijn stoffen die bij een verandering van de concentratie waterstofionen in het bloed de pH constant kunnen houden.
Zij zijn opgebouwd uit moleculen, die zowel H+-ionen kunnen
afgeven als opnemen. Wanneer het bloed te zuur dreigt te worden
fungeren zij als base. Wanneer de pH van het bloed dreigt te
stijgen, gegevens zij juist H+-ionen af. De belangrijkste buffers in
het bloed zijn fosfaat, bicarbonaat, plasma-eiwitten en hemoglobine;
– ademhaling; bij de uitademing wordt koolstofdioxide weer uit het
bloed verwijderd;
– urineproductie; een overschot aan H+-ionen kan met de urine geloosd
worden. De nieren zijn bovendien in staat om bij een acidose extra
bicarbonaat te vormen en aan het bloed af te geven. Dit bicarbonaat
bindt het overschot aan H+-ionen.
Intermezzo 1.2 Acidoses en alkaloses
Op basis van de oorzaak worden acidoses en alkalose ingedeeld
in respiratoire en non-respiratoire (‘metabole’) vormen.
Een respiratoire acidose komt voor bij patiënten met ademhalingsproblemen. Zij zijn niet meer in staat om alle CO2 uit te ademen.
De concentratie hiervan in het bloed zal stijgen en de pH zal dus
dalen. De patiënt zal dit proberen te compenseren door sneller
en dieper adem te halen en meer zuur via de nieren te lozen. Na
24 uur gaan de nieren bicarbonaat produceren. Deze base wordt
aan het bloed afgegeven waardoor de acidose verder wordt gecompenseerd.
Een respiratoire alkalose is meestal het gevolg van een te snelle
ademhaling (hyperventileren).
Een metabole acidose komt onder andere voor bij diabetici omdat
hun stofwisseling zogenaamde ketozuren vormt. Deze patiënten
hebben een opvallende snelle ademhaling om de pH te corrigeren.
Metabole alkaloses kunnen optreden bij langdurig braken of bij het
gebruik van bepaalde diuretica (‘plaspillen’). Het lichaam verliest dan te veel waterstofionen.
1.4
De fundamentele eenheid van het lichaam: de cel
Zoals al eerder is vermeld, vormt de cel de kleinste fundamentele
eenheid van leven, zowel wat bouw en structuur als wat werking en
functie betreft. Het is de kleinste levende bouwsteen van het menselijk lichaam. De leer van de cel wordt cytologie genoemd.
1
Cellen en weefsels
27
1.4.1
bouw en functie van cellen
De buitenste begrenzing van een menselijke cel wordt gevormd door
een celmembraan. Dit is in tegenstelling tot bij plantencellen en bacteriën. Deze laatste organismen leven vaak in een voor hen vijandige
omgeving. Als bescherming ligt er daarom om het celmembraan nog
een tweede laag: de celwand. De werking van antibiotica als penicilline is erop gebaseerd, dat deze middelen de celwand afbreken. Omdat deze bij menselijke cellen niet voorkomt zijn deze niet gevoelig
voor dit middel. Het binnenste van de cel wordt gevormd door een
waterige oplossing, het cytoplasma, met daarin een groot aantal bestanddelen, de zogenaamde organellen. Organellen zijn celstructuren
met een bepaalde functie: werkplaatsen van de cel. Het cytoplasma
zelf bestaat vooral uit water (75%) met daarin opgelost zouten,
eiwitten, koolhydraten en vetten. De eiwitten hebben onder andere
belangrijke enzymfuncties.
Hieronder volgt een bespreking van de bouw en functie van het
celmembraan en de belangrijkste organellen (afb. 1.1 en afb. 1.2).
celmembraan
actinefilamenten
ruw endoplasmatisch
reticulum
poriën
centriool met
microtubuli
peroxisoom
nucleolus
kern
vesikel
golgi-complex
glad endoplasmatisch
reticulum
lysosoom
mitochondriën
intermediaire
filamenten
Celmembraan
Het celmembraan wordt ook wel aangeduid als plasmamembraan. De
basisstructuur is een dubbele laag fosfolipiden (afb. 1.3). Fosfolipidenmoleculen hebben een vetoplosbaar en een wateroplosbaar deel.
De moleculen rangschikken zich zo, dat de vetoplosbare delen naar
elkaar toe wijzen en de wateroplosbare naar buiten, naar het cytoplasma en de vloeistof buiten de cel. Op deze manier vormt het
celmembraan een barrière voor wateroplosbare stoffen. Vetoplosbare
stoffen, zoals vetoplosbare hormonen, kunnen wel door het celmembraan diffunderen. Voor bepaalde stoffen als glucose, water en
aminozuren zijn specifieke carriers in het celmembraan aanwezig.
Deze carriers hebben een eiwitstructuur. Soms vormen deze eiwitten
simpelweg een kanaal door het membraan, soms werken zij als een
pomp. In dit laatste geval kunnen zij ten koste van energie stoffen
tegen de concentratie in vervoeren, dat wil zeggen van een lage naar
een hoge concentratie. Dan is er sprake van actief membraantransport.
Afbeelding 1.1 Schematische
voorstelling van een cel met daarin
de algemeen voorkomende
organellen.
28
Fysiologie en anatomie niveau 5
Afbeelding 1.2 Een cel,
elektronenmicroscopisch
(vergroting 20.0006).
1 celkern
2 kernmembraan
3 celmembraan
4 golgi-apparaat
5 glad endoplasmatisch reticulum
6 mitochondriën
7 celkern
8 lysosoom
9 ruw endoplasmatisch reticulum
1
2
3
4
5
6
7
8
9
Afbeelding 1.3 Structuur van het
celmembraan volgens het vloeibaar
mozaı̈ekmodel. De bimoleculaire
lipidenlaag bevat allerlei eiwitten.
extracellulair
glycoproteïne
glycocalix
glycolipide
integraal
eiwit
dubbele fosfolipidenlaag
cytoplasma
integraal eiwit
filamenten van cytoskelet
cholesterol
perifeer eiwit
intracellulair
Het leefmilieu van de cel is het waterige milieu rondom de lichaamscellen: het weefselvocht (andere benamingen daarvoor zijn:
interstitieel vocht, intern milieu, weefselvloeistof en intercellulaire
vloeistof ). Dit heeft vrijwel dezelfde samenstelling als bloedplasma en
varieert dus, bijvoorbeeld na een maaltijd. Door de selectieve opname
van voedingsstoffen is het milieu in de cel veel constanter.
Naast fosfolipiden en eiwitten vormt cholesterol de derde bouwsteen
van celmembranen. Cholesterol geeft stevigheid aan het celmembraan, vooral in membranen van zenuwcellen.
Het celmembraan bezit soms uitstulpingen: microvilli (borstelzoom)
(zie par. 10.14.2 en afb. 10.15 en 10.16). Deze vergroten het oppervlak
en daardoor het resorberend vermogen van de cel.
1
Cellen en weefsels
Kern
De kern (nucleus) is omgeven door het kernmembraan dat het kernplasma (nucleoplasma) omsluit. Het kernmembraan is een dubbelmembraan met poriën, waardoor direct contact mogelijk is tussen de
kern en het cytoplasma. In het kernplasma (protoplasma van de kern)
bevinden zich het chromatine en een of meer kernlichaampjes (nucleoli).
Het chromatine, bestaande uit eiwitten en DNA, is een fijnkorrelige
structuur, die zich gemakkelijk laat kleuren. Voorafgaand aan kerndeling ontstaan uit het chromatine de chromosomen. De celkern is de
drager van de aanleg voor de erfelijke eigenschappen: de genen.
Doordat de genen uiteindelijk bepalen welke eiwitten en dus ook
welke enzymen er in het cytoplasma ontstaan, vervult de kern in de cel
de functie van regulator van de levensverrichtingen, dat wil zeggen
van de stofwisseling in de organellen, bijvoorbeeld de synthese van
eiwitten (o.a. enzymen) en de verbranding.
De kern speelt ook een belangrijke rol bij de celdeling, doordat
celdeling wordt voorafgegaan door kerndeling.
Endoplasmatisch reticulum
Het endoplasmatisch reticulum (ER) is een gesloten netwerk (reticulum)
van holten en kanalen, gelegen in het cytoplasma. Het wordt begrensd door twee dicht tegen elkaar liggende membranen, die een
voortzetting zijn van het eveneens uit twee lagen bestaande kernmembraan. Wanneer zich aan de buitenzijde van het endoplasmatisch
reticulum ribosomen bevinden, wordt gesproken van ruw endoplasmatisch reticulum. De ribosomen komen ook vrij in het cytoplasma
voor. Het zijn bolvormige tot ovaalvormige structuren die betrokken
zijn bij de eiwitsynthese. Het inwendige kanalensysteem van het ER
dient voor het transport van eiwitten die op het oppervlak zijn gesynthetiseerd.
Glad endoplasmatisch reticulum, dat geen ribosomen bevat, speelt
een rol in de synthese van vetten en steroı̈den in de cel. Het vormt
tevens een overgang naar het golgi-apparaat.
Golgi-apparaat
Het golgi-apparaat (golgi-complex) is opgebouwd uit een groot aantal
door membranen omgeven holten. Het staat enerzijds in verbinding
met het endoplasmatisch reticulum en anderzijds met het celmembraan. In de blaasjes van het golgi-apparaat worden producten, met
name eiwitten, die zijn gemaakt op het endoplasmatisch reticulum,
bewerkt voor hun functie binnen de cel en buiten de cel, bijvoorbeeld
enzymen. De blaasjes (met de enzyminhoud) versmelten met het
celmembraan zodat de enzymen buiten de cel gebracht worden. Zo
komen bijvoorbeeld de enzymen van de pancreascellen naar buiten
om in de dunne darm het voedsel te verteren. Er is sprake van exocytose (zie par. 1.6.3). Behalve eiwitten wordt in het golgi-apparaat
ook slijm geproduceerd en vervolgens afgegeven.
29
30
Fysiologie en anatomie niveau 5
Lysosomen
Lysosomen zijn bolvormige organellen ontstaan uit het golgi-apparaat.
Onder normale omstandigheden beschikken ze over enzymen voor de
intracellulaire vertering van macromoleculen, stoffen met een hoge
molecuulmassa (eiwitten, vetten, polysachariden). Bij afwezigheid of
deficiëntie hiervan ontstaan er ophopingen van voornoemde macromoleculen, die bekend staan als lysosomale stapelingsziekten (zie
hoofdstuk 13).
Ze bezitten een aantal enzymen, die in staat zijn om normale celbestanddelen te splitsen, waardoor de cel wordt gelyseerd (opgelost). In
een levende cel bezitten de lysosomen aan de buitenzijde een beschermend membraan. Wanneer een micro-organisme door een cel
wordt gefagocyteerd (in een blaasje opgenomen) versmelt het membraan van het lysosoom met het membraan van het fagocytoseblaasje.
De inhoud van de lysosomen komt zo bij het micro-organisme, dat
door de lysosomale enzymen wordt afgebroken. Lysosomen komen
dan ook veel voor in leukocyten (witte bloedcellen), de cellen van het
afweersysteem (zie hoofdstuk 3).
Wanneer het membraan van het lysosoom wordt vernietigd door
bijvoorbeeld sterke zuren of gifstoffen, dan treedt onmiddellijk autolyse op, hetgeen neerkomt op zelfvernietiging van de cel. Bij donorbloed worden de leukocyten door centrifugeren verwijderd, waardoor
het bloed langer houdbaar is.
Er is ook een direct verband tussen het verouderingsproces van de cel
en het aantal lysosomen. Naarmate de cellen ouder worden, neemt
ook het aantal lysosomen toe.
Mitochondriën
Mitochondriën zijn bolvormige tot langgerekte organellen, opgebouwd
uit een dubbelmembraan waarvan het binnenste veel plooien (cristae)
bezit. In mitochondriën speelt zich de reactie af tussen zuurstof, dat
via de ademhaling is opgenomen, en de voedingstoffen, die na vertering uit het maag-darmkanaal zijn opgenomen, met als eindproducten koolstofdioxide en water (aerobe verbranding). Deze reactie
levert de energie voor de cel (zie par. 1.5). Daarom bevatten vooral
cellen, die veel energie verbruiken veel mitochondriën. Voorbeelden
zijn levercellen, hart- en spiercellen.
Behalve in de celkern komt ook in de mitochondriën DNA voor.
Omdat de mitochondriën in een bevruchte eicel van de moeder afkomstig zijn speelt dit DNA een belangrijke rol bij erfelijkheids- en
stamboomonderzoek.
Centrosoom
Het centrosoom speelt een belangrijke rol bij de celdeling, doordat het
de ‘polen’ vormt. Een centrosoom is opgebouwd uit twee centriolen.
Ciliën en flagellen
Ciliën en flagellen zijn celaanhangsels die bij veel cellen voorkomen.
Zo bevatten de epitheelcellen in de luchtpijp ciliën (trilhaartjes), die
1
Cellen en weefsels
zorgen voor het transport van slijm. Zaadcellen bevatten flagellen
(zweepdraden), die zorgen voor de voortbeweging van deze cellen.
1.5
Celmetabolisme: verbranding in de cel
De activiteiten in de hierboven genoemde organellen zijn bepalend
voor de cel als geheel en dus ook voor het menselijk lichaam als
totaliteit. De verschillende celactiviteiten worden uiteindelijk gereguleerd door de celkern. De benodigde energie wordt opgewekt in de cel
zelf in de speciaal daartoe bestemde organellen: de mitochondriën.
De mitochondriën worden dan ook aangeduid als de krachtcentrales
in de cel. Bij de verbranding wordt onderscheid gemaakt in aerobe en
anaerobe verbranding.
In alle cellen is de directe energiebron voor de celactiviteiten de stof
die bekend staat onder de naam ATP, voluit adenosinetrifosfaat. ATP
is een zeer energierijke verbinding waarmee activiteiten kunnen
worden uitgevoerd.
Bij de verbranding wordt de energie tijdelijk opgeslagen in de vorm
van de stof ATP volgens de reactie: ADP + fosfaat ? ATP
Wanneer deze energie nodig is voor een of andere vorm van arbeid
(mechanische arbeid in spieren, chemische arbeid voor het anabolisme of elektrische arbeid voor impulsgeleiding) verloopt deze reactie in omgekeerde richting, waarbij de opgeslagen energie weer vrijkomt. Het geproduceerde ADP wordt vervolgens weer ‘opgeslagen’ in
de mitochondriën.
1.5.1
aerobe verbrandingsprocessen
Zoals bekend verloopt de afbraak meestal met behulp van zuurstof:
aerobe oxidatie. Brandstoffen als glucose en vetzuren worden tijdens de
reactie met zuurstof in de cel omgezet in koolstofdioxide en water.
Hierbij komt energie vrij.
1.5.2
anaerobe verbrandingsprocessen
Onder omstandigheden waarbij er een tekort is aan zuurstof verloopt
de verbranding anaeroob. Deze levert veel minder energie dan de aerobe
verbranding. Zo levert de anaerobe verbranding van glucose slechts
twee moleculen ATP per molecuul glucose, terwijl aeroob de winst 36
ATP per molecuul glucose is. Het eindproduct van de anaerobe verbranding is melkzuur, dat uiteenvalt in lactaat en waterstofionen,
waardoor de omgeving zuur wordt. Dit is bijvoorbeeld het geval in
een (over)belaste spier of in situaties waarin de stroomsnelheid van
het bloed sterk is verminderd, zoals bij een shock. De concentratie
lactaat in het bloed is daarom een goede maat om de ernst van een
shock in te schatten.
1.5.3
koolhydraten, vetten en eiwitten
De meest bekende brandstoffen zijn koolhydraten (sacharose, zetmeel)
en lipiden (vetten). In het spijsverteringskanaal worden de disachariden, zoals sacharose (sucrose), en de polysachariden (zetmeel) afge-
31
32
Fysiologie en anatomie niveau 5
broken tot glucose, dat dan als brandstof beschikbaar is (zie hoofdstuk 10). In de lever- en spiercellen ligt altijd glycogeen in opslag, dat
ook afgebroken kan worden tot glucose voor het oxidatieproces.
Vetten zijn een uitstekende brandstof. Een gram vet levert tweemaal
zoveel ATP als een gram koolhydraat. Dit komt door het relatief grote
aantal waterstofatomen per molecuul vet. In de laatste stap van het
aerobe verbrandingsproces (terminale ademhalingsketen) koppelen
deze atomen zich namelijk met zuurstof waardoor water ontstaat en
er ATP vrijkomt.
In de stofwisseling van koolhydraten en vetten komt een gemeenschappelijk product voor, namelijk acetylco-enzym-A (acetyl-CoA). Via
deze stof leveren alle brandstoffen bij aerobe afbraak het onmisbare
ATP. Ook van veel aminozuren, de bouwstenen van eiwitten (hoofdstuk 10) is bekend dat ze via een aantal tussenstappen in acetyl-CoA
kunnen worden omgezet. Acetylco-enzym-A neemt een centrale
plaats in bij alle metabole processen die in een cel plaatsvinden.
1.6
Celmembraantransport
Er zijn twee mechanismen waarop transport van kleine moleculen
door membranen kan plaatsvinden: passief transport, waarbij geen
ATP vereist is, en actief transport, waarvoor ATP nodig is.
Het transport van grote moleculen en deeltjes vindt plaats door exocytose en endocytose.
1.6.1
passief transport
Voor passief transport is zoals gezegd geen ATP vereist. Hieronder
vallen: diffusie, osmose en filtratie.
Diffusie
Wanneer we een suikerklontje in een theeglas doen en we roeren niet,
dan zien we na verloop van tijd slierten van een dikke suikeroplossing
van de bodem van het glas naar boven trekken. Na lange tijd heeft de
suiker zich gelijkmatig door de thee verspreid. Wanneer iemand met
deodorant spuit ruiken we dat na een tijdje in het hele huis. Dit zijn
twee voorbeelden van diffusie. Onder diffusie wordt het verschijnsel
verstaan dat gassen, veel vloeistoffen (de zogenaamde mengbare
vloeistoffen) en oplossingen spontaan vermengen. Diffunderende
stoffen bewegen zich van plaatsen met een hoge concentratie naar
plaatsen met een lage concentratie, totdat de concentraties overal
gelijk zijn (Latijn: diffundere = zich verspreiden).
Diffusie wordt veroorzaakt door de beweging van de moleculen. De
drijvende kracht achter diffusie is het streven van de natuur om de
concentratie van stoffen overal gelijk te krijgen. Hoe groter de beweeglijkheid van de moleculen is, des te sneller verloopt de diffusie.
Het blijkt dan ook dat de diffusie bij gassen sneller verloopt dan bij
vloeistoffen.
De snelheid van een diffusieproces hangt af van een aantal factoren:
1
Cellen en weefsels
– temperatuur, naarmate de temperatuur hoger is neemt de snelheid
toe;
– molecuulmassa; kleine moleculen diffunderen sneller dan grote moleculen;
– concentratieverschil; hoe groter dit verschil des te sneller de diffusie;
het concentratieverschil wordt ook wel diffusiegradiënt genoemd;
– diffusieoppervlak; door een groot oppervlak kunnen per tijdseenheid
meer moleculen passeren;
– afstand, de dikte van het diffusiemembraan: hoe groter de afstand,
des te langzamer verloopt de diffusie.
De sterkte van de diffusiestroom wordt uitgedrukt in de wet van Fick,
waarbij concentratieverschil, membraanoppervlak en membraandikte
zijn betrokken (zie verder par. 4.3, waarin de diffusieprocessen van de
gassen zuurstof en koolstofdioxide worden besproken). Bij de ademhaling is sprake van een groot concentratieverschil (spanningsverschil) en een groot diffusieoppervlak gekoppeld aan een zeer dun
diffusiemembraan. Hierdoor zijn de voorwaarden vervuld om in een
kort tijdsbestek grote hoeveelheden zuurstof vanuit de alveoli (longblaasjes) naar het bloed over te brengen en omgekeerd zeer veel CO2
vanuit het bloed naar de alveoli. In de weefsels vinden eveneens
diffusieprocessen plaats. Overigens zijn er in de fysiologie maar weinig stoffen die vrij over een celmembraan kunnen diffunderen. Dit
zijn CO2, O2 en ureum (een afvalproduct van de eiwitstofwisseling).
In sommige gevallen wordt de diffusie vergemakkelijkt door speciale
carriers (’ondersteunende diffusie’, afb. 1.4). Dit zijn membraaneiwitten, die specifiek stoffen over een celmembraan kunnen transporteren. Een voorbeeld van een dergelijk carriertransport is het transport
van glucose over het celmembraan. De glucosecarrier bindt aan de
buitenzijde van de cel een molecuul glucose. Hierna ondergaat het
carriereiwit een vormverandering zodat het glucosemolecuul zich aan
de andere zijde van het membraan bevindt. Het glucosemolecuul
wordt losgelaten en de carrier neemt zijn oorspronkelijke vorm weer
aan, waarna het volgende glucosemolecuul kan worden overgebracht.
Het zal duidelijk zijn dat transport door middel van een carrier in
tegenstelling tot vrije diffusie aan een maximum is gebonden. Dit
maximum wordt bepaald door de snelheid waarmee de carrier van
vorm verandert. Carriers zijn over het algemeen zeer specifiek. Zo kan
de glucosecarrier geen andere suikers dan glucose over het celmembraan vervoeren. Deze carrier staat onder controle van het hormoon
insuline (zie hoofdstuk 7).
Veel carriers kunnen alleen functioneren wanneer zij twee deeltjes
tegelijk kunnen vervoeren. De bovenbeschreven glucosecarrier neemt
tegelijk met het glucosemolecuul ook een Na+-ion mee naar binnen.
De beweging van de deeltjes over het membraan kan ook tegengesteld
zijn. In de nieren komen carriers voor, die alleen functioneren als zij
een Na+-ion en een H+-ion in tegengestelde richting vervoeren. De
gevolgen hiervan zijn merkbaar in het geval van een acidose. De
nieren proberen deze te compenseren door extra H+-ionen uit te
33
34
Fysiologie en anatomie niveau 5
Afbeelding 1.4 Schematische
voorstelling van vrije diffusie (A)
en diffusie ondersteund door
membraaneiwitten (B) door het
plasmamembraan.
A vrije diffusie
B ondersteunde diffusie
extracellulaire
vloeistof
+
+
kanaaleiwit
hydrofiel
deeltje
+
+
transporteiwit
+
plasmamembraan
cytoplasma
+
+
hydrofoob
deeltje
ion
+
intracellulaire
vloeistof
scheiden. In plaats daarvan neemt de uitscheiding van natrium af,
waardoor de Na+-concentratie in het bloedplasma zal stijgen.
Osmose
Onder het begrip osmose wordt de diffusie van water door een semipermeabel (halfdoorlatend) membraan verstaan. Dit is een membraan
waarbij het oplosmiddel (water) wel kan passeren en de opgeloste
stof niet of slechts zeer langzaam. Een voorbeeld hiervan is te zien in
afbeelding 1.5. Een suikeroplossing wordt gescheiden van water door
een semipermeabel vlies. Aanvankelijk is daarin een geconcentreerde
suikeroplossing aanwezig. Doordat de natuur altijd streeft naar
evenwicht zal in dit geval, doordat suiker het membraan niet kan
passeren, water zich gaan verplaatsen in de richting van de suikeroplossing. De suikeroplossing oefent dus een aanzuigkracht uit op
het omringende water (osmos = aandrang). De aanzuigkracht wordt
osmotische druk genoemd. De grootte van de osmotische druk wordt
bepaald door de concentratie van deeltjes die niet over het semipermeabele membraan kunnen diffunderen.
Afbeelding 1.5 Osmose.
niet
geconcentreerde
oplossing
geconcentreerde
oplossing
H2O
semipermeabele
membraan
1
Cellen en weefsels
Het verschijnsel osmose is ook op een andere manier te verklaren.
Hiertoe dient het begrip waterconcentratie gehanteerd te worden. In een
oplossing is er dan sprake van de concentratie van de opgeloste stof
en van de waterconcentratie. In een geconcentreerde oplossing is er
dus een hoge concentratie van de opgeloste stof en een ‘geringe’
waterconcentratie. Wanneer een geconcentreerde suikeroplossing
door een semipermeabel vlies is gescheiden van zuiver water (de
waterconcentratie is dan 100%) zal water naar de suikeroplossing
diffunderen omdat daarin de waterconcentratie geringer is. Op deze
wijze is het duidelijker dat osmose een kwestie is van diffusie, namelijk diffusie van water.
Osmose speelt een belangrijke rol bij de cellen van alle organismen
doordat membranen semipermeabel zijn. Deze doorlaatbaarheid
verschilt voor verschillende membranen. Het membraan van de wand
van de capillairen (haarvaten) is voor alle in het bloed opgeloste
stoffen doorlaatbaar, met uitzondering van eiwitten. Natrium, kalium
en glucose kunnen dus vrij het bloed uit diffunderen, eiwitten niet. De
eiwitconcentratie bepaalt daarom de osmotische druk van het bloed.
Omdat eiwitten grote moleculen (colloı̈den) zijn wordt deze druk vaak
aangeduid met de colloı̈d-osmotische druk (COD). Tegenwoordig wordt de
COD ook vaak oncotische druk (Grieks: oncos = zwelling) genoemd
vanwege de relatie met oedeemvorming.
Voor het celmembraan ligt de situatie anders. Deze is slechts voor
zeer weinig stoffen doorlaatbaar, meestal alleen wanneer er voor die
stoffen (bijv. glucose) een transportsysteem is aangelegd. Het celmembraan is bijvoorbeeld niet doorlaatbaar voor natrium, kalium en
chloride. De osmotische druk over een celmembraan wordt daarom
vooral bepaald door de zoutconcentratie. Dit wordt de kristalloı̈d-osmotische druk genoemd. Samenvattend: transport van water over het
membraan van de capillairen wordt bepaald door het verschil in
eiwitconcentratie binnen en buiten de bloedbaan: de colloı̈d-osmotische druk. Transport van water over het celmembraan wordt vooral
bepaald door een verschil in zoutconcentratie binnen en buiten de
cel: de kristalloı̈d-osmotische druk.
Intermezzo 1.3 Bepaling van de osmolariteit van het
bloedplasma
De colloı̈d-osmotische waarde van het bloedplasma volgt rechtstreeks uit de concentratie totaal eiwit in het bloedplasma. Om
de kristalloı̈d-osmotische waarde te berekenen zou eigenlijk de
concentratie van alle zouten in het bloedplasma bepaald moeten
worden. Dit is natuurlijk ondoenlijk. Daarom worden in de
praktijk allerlei vereenvoudigingen gehanteerd. Veel gebruikte
formules zijn:
kristalloı̈d-osmotische waarde = 2 [Na+ + K+]
Omdat tegenover ieder Na+- en K+-ion per definitie een negatief
deeltje moet voorkomen, wordt de concentratie van deze ionen
met twee vermenigvuldigd.
35
36
Fysiologie en anatomie niveau 5
Iets nauwkeuriger is de volgende berekening:
kristalloı̈d-osmotische waarde = 2 [Na+ + K+] + glucose + ureum
In het bloed treden osmotische verschijnselen op, onder andere bij de
erytrocyten (rode bloedcellen). Wanneer het bloedplasma een hogere
zoutconcentratie bezit (en dus een lagere waterconcentratie) dan de
erytrocyten, zal er diffusie van water plaatsvinden vanuit de erytrocyten naar het bloedplasma (afb. 1.6a). In de erytrocyten is in dat geval
de waterconcentratie namelijk hoger dan in het bloedplasma. In dit
geval is het bloedplasma hyperosmotisch ten opzichte van de bloedcellen. De term hyperosmotisch verdient de voorkeur boven de gebruikelijke term hypertonisch omdat de laatste wordt gehanteerd voor
afzonderlijke deeltjes. Hetzelfde geldt voor de termen isotonisch en
hypotonisch, die daarom nu bij voorkeur worden aangeduid respectievelijk als iso-osmotisch en hypo-osmotisch.
Wanneer het bloedplasma minder opgeloste stoffen bevat dan de
erytrocyten is het bloedplasma hypo-osmotisch (Grieks: hypo = onder)
ten opzichte van de erytrocyten. De consequentie hiervan is dat er
diffusie van water zal plaatsvinden vanuit het bloedplasma naar de
erytrocyten. Immers, in dit geval is de waterconcentratie in het
bloedplasma hoger dan in de erytrocyten. Het gevolg is dat de erytrocyten zwellen en ten slotte barsten. Dit verschijnsel wordt hemolyse genoemd (afb. 1.6b). Wanneer twee oplossingen dezelfde osmotische waarde bezitten worden ze iso-osmotisch (Grieks: isos = gelijk)
ten opzichte van elkaar genoemd. Onder normale omstandigheden is
het bloedplasma dan ook altijd iso-osmotisch ten opzichte van de
vloeistof in de erytrocyten.
Afbeelding 1.6 a Erytrocyt in een
hyperosmotisch milieu. De cel
krijgt een doornappelvorm. b
Erytrocyt in een hypo-osmotisch
milieu. Er treedt hemolyse op.
a
b
1
Cellen en weefsels
Intermezzo 1.4 Infusievloeistoffen
Wanneer een patiënt ‘vocht’ moet worden toegediend wordt
uiteraard ook gebruikgemaakt van een iso-osmotische oplossing. Een oplossing van 0,9% NaCl heeft dezelfde osmotische
waarde als het bloedplasma. Deze veel gebruikte iso-osmotische
oplossing wordt fysiologisch zoutoplossing (‘fysiologisch zout’)
genoemd, omdat door toediening van deze vloeistof de normale
fysiologie van de erytrocyten intact blijft.
Een iso-osmotische glucoseoplossing zal een andere concentratie moeten hebben dan een iso-osmotische oplossing van keukenzout. Op de eerste plaats splitst NaCl (keukenzout) wanneer
het wordt opgelost in water in twee deeltjes, te weten een natriumion en een chloride-ion. De formule van glucose is
C6H12O6, dus een glucosemolecuul is opgebouwd uit in totaal 24
atomen (6 koolstofatomen, 12 waterstofatomen en 6 zuurstofatomen). Bij het oplossen in water blijven deze atomen echter
aan elkaar gebonden. Een oplossing van 1 mol keukenzout per
liter levert om die reden tweemaal zoveel deeltjes als een oplossing van 1 mol glucose per liter. Omdat het aantal deeltjes per
liter de osmolariteit bepaalt is de osmolariteit van 1 mol keukenzout per liter daarom tweemaal zo groot als die van een
oplossing van 1 mol glucose per liter. Wanneer de concentratie
in grammen per liter of in procenten wordt uitgedrukt, speelt er
nog een tweede factor mee. Een glucosemolecuul is ongeveer
driemaal zo zwaar als een keukenzoutmolecuul. Voor een gelijk
aantal deeltjes is dus driemaal zoveel glucose als keukenzout
nodig.
Wanneer beide bovenstaande factoren worden gecombineerd,
luidt de conclusie dat er voor een iso-osmotische glucoseoplossing ongeveer 6 maal zoveel stof nodig is als voor een
fysiologische zoutoplossing. Dat komt overeen met een oplossing van glucose van ruim 5% (50 gram glucose per liter). Voor
infusievloeistoffen wordt meestal 5% glucose gebruikt. In dit
verband zal het ook duidelijk zijn dat wanneer de cellen glucose
opslaan in de vorm van glycogeen (zie hoofdstuk 10) het osmotisch effect van een glucose-infuus sterk wordt gereduceerd.
Infusievloeistoffen bevatten naast NaCl of glucose vaak nog
andere bestanddelen. Zo zal er in het geval van een patiënt met
een acidose een base, meestal bicarbonaat, aan het infuus worden toegevoegd. Bij een alkalose wordt er een zuur (bijvoorbeeld
zoutzuur) aan het infuus toegevoegd. Om de iso-osmolariteit
van het infuus te handhaven, moet de concentratie van deze
toevoegingen op de concentratie zout of glucose in mindering
worden gebracht.
Bij een patiënt, die een groot bloedvolume heeft verloren wordt
soms een grootmoleculaire stof (albumine of dextraan, een soort
zetmeel) aan het infuus toegevoegd. Hiermee wordt de colloı̈d-
37
38
Fysiologie en anatomie niveau 5
osmotische druk van het bloed verhoogd, waardoor een groter
deel van de infusievloeistof in de bloedbaan zal blijven.
Filtratie
Filtratie is het proces, waarbij water met opgeloste stoffen zich over
een wand verplaatst. De drijvende kracht achter filtratie is de hydrostatische druk (druk die door het water wordt uitgeoefend). Filtratie
speelt een belangrijke rol bij het uittreden van bloedplasma met voedingsstoffen uit de capillairen (zie hoofdstuk 2) en bij de vorming van
urine (zie hoofdstuk 5). Het proces zal in die hoofdstukken verder
worden toegelicht.
1.6.2
actief transport
Voor actief transport is ATP vereist. Het mechanisme achter actief
transport vertoont veel overeenkomsten met het beschreven carriertransport. Ook nu is er sprake van een eiwit in het celmembraan, dat
vormveranderingen ondergaat, waarvoor in dit geval ATP nodig is.
Een bijzondere eigenschap van actief transport is dat hiermee deeltjes
ook van een lage naar een hoge concentratie kunnen worden vervoerd. Het bekendste voorbeeld van actief transport is de natrium/
kaliumpomp (Na/K-pomp). Deze pomp vervoert tegelijkertijd natriumionen vanuit de cel naar het interstitium en kaliumionen vanuit het
interstitium de cel in. In beide gevallen is dat van een plaats met een
lage concentratie naar een plaats met een hoge concentratie.
De Na/K-pomp speelt een belangrijke rol bij de impulsgeleiding in de
hartspiercellen (zie hoofdstuk 2) en de zenuwcellen (zie hoofdstuk 6).
1.6.3
endocytose en exocytose
Endocytose en exocytose worden samen ook wel aangeduid met
blaasjestransport.
Bij endocytose omsluit het celmembraan een deel van de vloeistof in het
interstitium met de daarin opgeloste stoffen. De aldus gevormde
blaasjes (vesikels) worden als kleine organellen in de cel opgenomen.
Daarna kunnen zij bijvoorbeeld versmelten met de membranen van de
lysosomen, waardoor de inhoud van deze twee organellen versmelt.
Levercellen bijvoorbeeld zijn door endocytose in staat te veel cholesterol uit het bloed te verwijderen en in de lysosomen op te nemen.
Cholesterol wordt vervolgens in de lysosomen afgebroken.
Een bijzondere vorm van endocytose is fagocytose (zie hoofdstuk 3).
Hierbij worden vaste deeltjes als bacteriën in de cel opgenomen.
Wanneer vloeibaar materiaal in een cel wordt opgenomen wordt dat
pinocytose genoemd (Grieks: pinein = drinken).
Omgekeerd kunnen in de cel gevormde stoffen worden verpakt in een
blaasje van membraanmateriaal. Als het membraan van dit blaasje
vervolgens versmelt met het celmembraan van de cel kan deze stof
naar het interstitium worden afgegeven. Dit heet exocytose (afb. 1.7 en
afb. 1.8). Het genoemde golgi-apparaat speelt hierbij een belangrijke
rol.
1
Cellen en weefsels
39
glad
dubbele
endoplasmatisch
kernmembraan
reticulum
ruw
endoplasmatisch
golgi-complex
reticulum
celmembraan
Afbeelding 1.7 Schematische
voorstelling van de vorming van
een eiwit en afgifte door middel
van exocytose.
porie
kern
exocytose
mRNAtransport
eiwittransport
vesikelvorming
endocytose
eiwitsynthese
op te nemen deeltjes
1
2
3
golgi-complex
4
plasmamembraan
lysosoom
transcytose
synthese
plasmamembraan
exocytose
De combinatie van endocytose en exocytose komt onder andere voor
in de darmwand. Dit verschijnsel wordt wel transcytose genoemd.
Stoffen uit het voedsel worden dan door middel van endocytose in de
cellen van de darmwand opgenomen en daarna door exocytose afgegeven aan de bloedvaten of de lymfevaten (zie hoofdstuk 10).
1.7
Celdeling
Het menselijk lichaam ontwikkelt zich door deling en groei vanuit de
zygote (bevruchte eicel). Bij deling van een cel ontstaan dochtercellen
die vrijwel identiek zijn aan de moedercel. Iedere celdeling wordt
voorafgegaan door een kerndeling.
Bij zich delende cellen wordt een celcyclus onderscheiden die in twee
stadia wordt onderverdeeld, namelijk de mitose en de interfase.
Onder mitose wordt de celdeling verstaan, waarbij uit één cel twee
genetisch identieke dochtercellen ontstaan die hetzelfde aantal chromosomen bezitten als de moedercel. De mitose, ook wel somatische
celdeling of ‘gewone’ celdeling genoemd, duurt afhankelijk van het
celtype en milieuomstandigheden 1-2 uur.
Afbeelding 1.8 Schematische
voorstelling van import, export en
transcellulair transport. In het
laatste geval bestaat er een
combinatie van endo- en
exocytose.
40
Fysiologie en anatomie niveau 5
De interfase is het stadium tussen twee op elkaar volgende mitosen. De
chromosomen zijn dan langgerekte dunne draden: het chromatine.
De duur van de interfase is erg variabel, afhankelijk van het celtype en
de milieu-omstandigheden. Hij duurt in ieder geval vele malen langer
dan de mitose zelf, bijvoorbeeld dertig uur. Tijdens de interfase vinden achtereenvolgens plaats: celgroei, verdubbeling (replicatie) van
de chromosomen (zie ook hoofdstuk 12) en een directe voorbereiding
op de mitose. Tijdens de interfase zijn de chromosomen microscopisch niet waarneembaar in tegenstelling tot de chromosomen in de
mitose (afb. 1.9).
Afbeelding 1.9 Microfoto
(vergroting 55006) van de 46
chromosomen van de cellen van
een vrouw (metafase).
1.7.1
mitose
De mitose wordt globaal verdeeld in vier fasen (afb. 1.10):
– profase; voorafgaand aan de profase hebben de draden van het
chromatine zich verdubbeld waarna chromosomen ontstaan.
Hierin zijn de langgerekte chromatinedraden gespiraliseerd (opgerold), waardoor zij kort en dik worden en onder de microscoop
te zien zijn. De beide centrosomen (centriolen) komen verder van
elkaar te liggen en begeven zich naar de polen. Het kernmembraan
en de kernlichaampjes, die tijdens de interfase nog wel zichtbaar
zijn, verdwijnen;
– metafase; de chromosomen zijn inmiddels nog korter en dikker geworden. In het cytoplasma is een spoelvormige figuur van fijne
draden ontstaan, de zogenaamde kernspoel. De chromosomen zijn
inmiddels in het midden van de cel, het equatorvlak, aangekomen.
De beide chromatiden van een chromosoom zitten nog aan elkaar
vast door middel van het centromeer, een klein, nog ongedeeld
lichaampje. Een gedeelte van de spoeldraden (de trekdraden) is er
aan bevestigd;
1
Cellen en weefsels
41
– anafase; de centromeren delen zich en de twee chromatiden van
ieder chromosoom worden door de trekdraden naar de polen (centrosomen) getrokken. De zelfstandige chromatiden worden vanaf dit
moment weer chromosomen genoemd;
– telofase; wanneer de chromosomen bij de polen zijn aangekomen,
gaan ze zich despiraliseren, waarbij ze weer lang, dun en steeds
minder duidelijk worden. Rondom iedere pool wordt een kernmembraan gevormd en de kernlichaampjes worden weer zichtbaar.
Er volgt dan een insnoering van het celmembraan ter hoogte van
het equatorvlak, waarbij het cytoplasma over de twee nieuwe cellen
wordt verdeeld.
Na de telofase gaan veel cellen zich specialiseren waardoor ze nooit
meer aan een volgende celdeling toekomen.
celmembraan
kernmembraan
kernlichaampje
centromeer
chromatine
chromosoom
(twee chromatiden)
centriolen
interfase
profase
pool
spoelfiguur
metafase
anafase
telofase
interfase
Afbeelding 1.10 Schema van de mitose en microfoto’s van profase, metafase en anafase.
Bij witte bloedcellen is een celdeling waargenomen waarbij het cytoplasma en de kern snel in tweeën worden gedeeld zonder dat er
chromosomen zichtbaar worden. De cel strekt zich hierbij uit tot een
halter die in het midden splijt. Dit wordt amitose genoemd. Ook degenererende of abnormale weefselcellen (tumorcellen) kunnen zich
42
Fysiologie en anatomie niveau 5
soms amitotisch delen. Bij de amitose treedt waarschijnlijk geen
splitsing van chromosomen op.
1.7.2
meiose of reductiedeling
In de meeste cellen van het menselijk lichaam komen 23 paar chromosomen voor (afb. 1.11); dergelijke cellen worden diploı̈d genoemd.
Dit is ook het geval bij de cellen waaruit de voortplantingscellen
(gameten) ontstaan. De laatste celdelingen bij de vorming van deze
gameten geschieden anders dan bij de mitose. Deze worden meiose of
reductiedeling genoemd omdat bij deze celdelingen het aantal chromosomen per nieuwgevormde kern met de helft wordt verminderd.
De gameten hebben van ieder chromosomenpaar slechts de helft.
Deze cellen heten haploı̈d, wat wordt aangeduid met de letter n (= 23).
Wanneer de mannelijke en de vrouwelijke gameet met elkaar versmelten, zal de kern van zygote weer het normale aantal chromosomen bevatten: diploı̈d (2n) = 23 paar. Door de meiose blijft dus het
aantal chromosomen bij de geslachtelijke voortplanting constant.
Afbeelding 1.11 a Karyogram
(‘chromosomenportret’) van een
vrouw.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Y
X
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Y
X
Afbeelding 1.11 b Karyogram van
een man.
De meiose omvat twee delingsprocessen, kortweg meiose I en meiose
II genoemd (afb. 1.12).
Meiose I
Hierbij worden de volgende fasen onderscheiden:
– profase I; deze duurt veel langer dan de profase in de mitose. De
chromosomen bestaan aanvankelijk ieder nog uit twee chromatiden zoals bij de mitose. De homologe chromosomen (de twee op elkaar
gelijkende chromosomen van één paar) gaan naar elkaar toe en
1
Cellen en weefsels
43
Afbeelding 1.11 c Karyogram van
een downpatiënte.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Y
X
celmembraan
centromeer
chromatide
profase l
metafase l
anafase l
telofase l
anafase ll
telofase ll
Afbeelding 1.12 Meiose. In de afbeeldingen zijn voor de duidelijkheid slechts drie paar chromosomen weergegeven.
Profase II en metafase II zijn niet afgebeeld.
gaan ‘paren’, waarbij ze korter en dikker worden. Op dat moment
vindt er uitwisseling van genen (stukjes DNA) plaats tussen deze
twee homologe chromosomen. Hierdoor ontstaat steeds nieuwe
combinaties van eigenschappen, waardoor het aantal verschillende
nakomelingen enorm wordt uitgebreid. Dit proces wordt ‘crossingover’ genoemd. Daarna beginnen de homologe chromosomen
elkaar af te stoten; op bepaalde plaatsen blijven ze echter nog bij
elkaar: de chiasmata. De chromosomen worden nog korter; kernmembraan en kernlichaampjes verdwijnen en de kernspoel wordt
zichtbaar;
– metafase I; de chromosomen rangschikken zich in het equatorvlak;
de centromeren delen zich niet;
– anafase I; de scheiding van de homologe chromosomen wordt nu
voltooid; de chromosomen, ieder bestaande uit twee chromatiden,
begeven zich naar de polen;
– telofase I; de chromosomen komen bij de polen. Er wordt een
kernmembraan gevormd en tevens een celmembraan, waardoor er
twee cellen zijn ontstaan.
De kernen hebben dus ieder slechts de helft van het aantal chromo-
44
Fysiologie en anatomie niveau 5
somen van de moedercel waaruit ze zijn ontstaan. In de meiose I vindt
dus de reductie van het aantal chromosomen plaats.
Intermezzo 1.5 Het syndroom van Down
Ongeveer 95% van de mensen met het syndroom van Down heeft
een trisomie 21. Dat wil zeggen dat er van het chromosoom 21
drie in plaats van twee exemplaren in de lichaamscellen voorkomen. Dit is het gevolg van non-disjunctie tijdens (meestal de
eerste) meiotische deling. Hierbij gaan tijdens de vorming van de
ei- of zaadcel de twee exemplaren van chromosomenpaar 21 niet
uit elkaar, maar gaan beide chromosomen naar één dochtercel,
die dan 24 chromosomen heeft. De lichaamscellen, die na de
bevruchting uit de bevruchte eicel ontstaan hebben dan steeds
drie exemplaren van chromosoom 21 (zie afb. 1.11c). De eigenlijke oorzaak van de non-disjunctie is niet bekend, maar de
afwijking komt vaker voor in eicellen bij vrouwen ouder dan 37
jaar. Waarschijnlijk speelt ook de leeftijd van de vader een rol. De
non-disjunctie vindt in 80% van de gevallen plaats tijdens de
oögenese (vorming van eicellen) en in ongeveer 20% tijdens de
spermatogenese (vorming van zaadcellen).
Meiose II
Deze deling kan beschouwd worden als een gewone deling (mitose)
maar dan uitgaande van een cel met half zoveel chromososmen als
normaal. Er wordt een nieuwe spoelfiguur gevormd die loodrecht op
de eerste staat. De centromeren delen zich nu, waardoor de chromatiden, die nu weer chromosomen genoemd worden, zich naar de
polen begeven. Na afloop van meiose I en II zijn er vier kernen ontstaan, de zogenaamde tetradefiguur. Iedere kern is haploı̈d. Als het
cytoplasma zich heeft gedeeld zijn er, na differentiatie, vier individuele gameten ontstaan. Bij de spermatogenese (vorming van zaadcellen) levert iedere moedercel vier functionele zaadcellen (zie afb.
12.11). Bij de oögenese (vorming van eicellen) blijft er echter slechts één
functionele eicel over die zeer veel cytoplasma (reservevoedsel) bevat.
Gedurende de meiose I krijgt één van de dochterkernen vrijwel al het
cytoplasma, terwijl de andere kern als zogenaamd poollichaampje
degenereert. Gedurende de meiose II wordt opnieuw een poollichaampje gevormd, waardoor uiteindelijk één grote eicel overblijft (zie
afb. 12.3). Bij de vrouw begint de eerste meiotische deling al voor de
geboorte. Deze stopt echter na de profase. Deze wordt pas weer vervolgd na de puberteit, waarbij tijdens iedere cyclus een of enkele
oöcyten meiose I afmaken. Deze wordt dan direct gevolgd door
meiose II. Bij de man start de meiose pas bij de puberteit, deze gaat
daarna het gehele leven door.
In afbeelding 1.13 is een schematisch overzicht van de mitose en de
meiose weergegeven.
Bij de conceptie ontstaat de bevruchte eicel (zygote). Deze is te be-
1
Cellen en weefsels
45
schouwen als de eerste moedercel en ontwikkelt zich door een reeks
klievingsdelingen tot een pre-embryo van 64 cellen. Hieruit ontstaat
onder andere een embryoblast van enkele cellen. Deze vormt de oorsprong van de embryonale stamcellen.
2n
2n
2n
mitose
n
n
n
n
n
n
2n
meiose l
meiose ll
Intermezzo 1.6 Stamcellen
Stamcellen zijn cellen die zich onbeperkt kunnen delen. Zoals
eerder vermeld ontstaan alle cellen door deling uit een moedercel. De eerste moedercel is de zygote. Deze cel is totipotent. De
eerste uren na de bevruchting deelt deze cel zich nog een aantal
keren, waardoor meerdere totipotente stamcellen ontstaan, dat
wil zeggen dat elk van deze cellen de mogelijkheid heeft om uit
te groeien tot een foetus.
Na ongeveer vier dagen ontstaat de blastocyste (kiemblaasje),
een met vocht gevuld blaasje. Uit de buitenste laag hiervan ontwikkelt zich onder andere de placenta. De binnenste celmassa is
pluripotent, dat wil zeggen dat uit deze cellen zich vele andere
verschillende soorten cellen kunnen ontwikkelen, echter niet
alle celtypen die nodig zijn voor de ontwikkeling van een foetus.
De pluripotente stamcellen specialiseren zich vervolgens tot
multipotente stamcellen. Op die manier ontstaan uit de multipotente hemopoëtische stamcellen, de erytrocyten, trombocyten
en leukocyten (rode bloedcellen, bloedplaatjes en witte bloedcellen).
Er worden drie soorten celpopulaties onderscheiden:
– statische celpopulaties; de cellen hiervan kunnen zich niet meer
delen; wanneer deze weefsels beschadigd worden kunnen ze
slechts hersteld worden door vervanging van deze cellen door
andere typen cellen, zoals bindweefselcellen. Voorbeelden van
deze populaties zijn cellen van zenuw- en spierweefsel;
Afbeelding 1.13 Schematisch
overzicht van mitose en meiose.
Mitose: uitgaande van een
diploı̈de cel (2n) ontstaan er twee
diploı̈de cellen. Meiose: bij de
meiose I wordt het aantal
chromosomen gereduceerd tot de
helft (n), waarna de meiose II
volgt, te vergelijken met een
mitose: het aantal chromosomen
blijft gelijk. Het eindresultaat is 4
haploı̈de gameten (voortplantingscellen).
46
Fysiologie en anatomie niveau 5
– stabiele celpopulaties; deze vormen bijvoorbeeld het hoofdbestanddeel van de lever; de cellen delen zich wanneer vervanging nodig is na beschadiging of ziekte;
– vernieuwende celpopulaties; deze zijn met name aanwezig in de
huid, het darmslijmvlies en bloedvormende weefsels; deze
cellen blijven zich voortdurend delen omdat ze steeds vervangen moeten worden wegens slijtage van de oppervlakkige
laag of als gevolg van een korte levensduur.
In de vernieuwende populaties zijn de moedercellen, stamcellen
geheten, minder ver ontwikkeld dan de dochtercellen. Deze
dochtercellen delen zich na rijping niet meer, maar de stamcellen kunnen zich onbeperkt delen. Stamcellen worden gekenmerkt door het feit dat wanneer ze zich delen ten minste één van
de twee ontstane dochtercellen hetzelfde primitieve karakter
heeft als dat van de oorspronkelijke oudercel, terwijl de andere
dochtercel een meer gespecialiseerd celtype kan worden. Primitief wil in dit verband zeggen dat ze aan het begin van de ontwikkeling staan. Stamcellen zijn dus zowel in staat zichzelf te
vermeerderen en zo meer stamcellen te produceren als dochtercellen te produceren die zich verder kunnen specialiseren tot
bijvoorbeeld bloed-, spier- of hersencellen.
Humane pluripotente stamcellen kunnen geı̈soleerd worden uit
de binnenste celmassa van de eerder genoemde blastocyste (een
vroeg ontwikkelingsstadium van het embryo). Deze embryonale
stamcellen worden gewonnen uit pre-implantatie-embryo’s, de
zogenaamde restembryo’s, die overgebleven zijn bij de ivf-behandeling (in-vitrofertilisatie of ‘reageerbuisbevruchting’).
De Embryowet biedt in Nederland de mogelijkheid om nieuwe
embryonale stamcellijnen te isoleren uit bestaande restembryo’s
die overblijven na ivf. Het is niet toegestaan om embryo‘s speciaal voor dit doel te creëren.
Pluripotente stamcellen kunnen ook verkregen worden door
somatische celkerntransplantatie. Uit een eicel wordt de kern
verwijderd. Vervolgens wordt een somatische cel bij de eicel gebracht en worden de twee gefuseerd. Na de fusie ontstaat een
totipotente stamcel. Na een aantal delingscycli ontstaat de blastocyste. In de binnenste celmassa bevinden zich de pluripotente
stamcellen. Uit proefdieronderzoek is gebleken dat het embryo,
dat uit een dergelijke celkernvervanging ontstaat, vaak abnormaal is omdat de kern van een bevruchte eicel verschilt van een
ingebrachte kern van een somatische cel.
Embryo’s die na celkernvervanging ontstaan, mogen volgens de
Embryowet niet in de uterus teruggeplaatst worden wegens de
grote kans op kinderen met ernstige aangeboren afwijkingen.
1
Cellen en weefsels
Onderzoek met humane pluripotente stamcellen heeft geleid tot
de mogelijkheid van celtherapie. Veel aandoeningen zijn het gevolg van een verstoorde celfunctie of beschadigde weefsels.
Pluripotente stamcellen kunnen, als ze aangezet worden tot
specifieke celspecialisatie, een bron zijn van de te vervangen
cellen bij bijvoorbeeld de ziekte van Parkinson, Alzheimer, diabetes mellitus, dwarslaesie en hartziekten.
Onderzoek van de laatste jaren op het gebied van stamcellen
heeft aangetoond dat het mogelijk is om nieuwe hartspiercellen
(cardiomyocyten) en bloedvatcellen (endotheelcellen en gladde
spiercellen) te verkrijgen uit stamcellen die uit het beenmerg
afkomstig zijn. Bij experimenten is het gelukt om stamcellen te
laten ontwikkelen tot cardiomyocyten.
Proefdieronderzoek heeft aangetoond dat gezonde hartspiercellen getransplanteerd in het hart met succes het hartweefsel vervangen.
Bij patiënten met de ziekte van Parkinson is celtherapie voor het
eerst toegepast. De getransplanteerde zenuwcellen zijn afkomstig van menselijke foetussen. Een groot probleem hierbij is dat
er vijf à tien foetussen nodig zijn om te beschikken over voldoende cellen voor één enkele transplantatie. De foetussen zijn
afkomstig van abortus en slechts in zeer beperkte mate beschikbaar. Sinds 1996 wordt er ook onderzoek gedaan met foetale varkenscellen (xenotransplantatie; xenos = vreemd). Maar
ook hierbij zijn de nodige bedenkingen.
Recent onderzoek heeft aangetoond dat nieuwe zenuwcellen
ook in volwassen hersenen worden aangemaakt. Deze nieuwvorming is beperkt tot bepaalde plaatsen en bescheiden van
omvang. Voor het herstel van grotere beschadigingen lijkt dit
proces onvoldoende. Er zijn drie plaatsen bekend waar nieuwe
zenuwcellen worden gemaakt: in het reukorgaan, in de reukkern
van de hersenen en in de hippocampus (gelegen tussen de grote
hersenen en de hersenstam; zie par. 6.14.3). Het is gelukt om
voorlopercellen uit het neusslijmvlies te kweken, waardoor de
mogelijkheid ontstaat om grote hoeveelheden zenuwcellen te
produceren.
1.8
Celcyclus
In de voorafgaande paragrafen is beschreven hoe cellen zich kunnen
delen en hoe cellen zich kunnen specialiseren tot een groot aantal
verschillende typen. Afhankelijk van het type cel varieert de levenscyclus van een cel van enkele dagen tot levenslang.
47
48
Fysiologie en anatomie niveau 5
1.8.1
celdood
Het leven van een cel kan op twee manieren eindigen, namelijk door
beschadiging en van nature. Cellen kunnen beschadigd worden door
agentia (invloeden) die de cellen rechtstreeks aantasten, bijvoorbeeld
micro-organismen (zoals bacteriën en virussen), toxinen (gifstoffen)
en chemisch actieve stoffen. Ook een indirecte inwerking, zoals een
tekort aan zuurstof of voedingsstoffen en een ophoping van afvalstoffen, kan een vervroegde celdood veroorzaken. Dit wordt necrose
(celversterf ) genoemd. Een cel kan ook uit zichzelf degenereren en
ten slotte afsterven. Dit gebeurt onder invloed van bepaalde genen,
wanneer een vermindering van het aantal cellen gewenst is. Dit heet
geprogrammeerde celdood of apoptose. Elke cel heeft in de mitochondriën een aantal eiwitten die een pre-apoptotische werking hebben. Wanneer de cel voldoende stimulerende signalen van andere
cellen krijgt blijven deze eiwitten inactief. Wanneer deze signalen
verdwijnen treedt een aantal reacties op. Het begint met het omzetten
van de pre-apoptotische eiwitten tot eiwitsplitsende enzymen. Deze
enzymen tasten verschillende celorganellen aan, waardoor de cel uiteenvalt in brokstukjes (apoptic bodies). Deze worden door andere
cellen gefagocyteerd en door lysosomen verteerd. Hierbij komen in
tegenstelling tot bij necrose geen afvalstoffen vrij, waardoor er ook
geen sprake kan zijn van ontstekingsreacties. Apoptose is een normaal verschijnsel, waarbij overbodige cellen worden opgeruimd.
1.8.2
groei
Bij de celcyclus treedt een aantal opeenvolgende processen op dat
goed geprogrammeerd moet worden. Het betreffende weefsel of orgaan moet zijn normale grootte en vorm behouden. Wanneer een
orgaan in volume afneemt wordt gesproken van hypo- of atrofie.
Wanneer een orgaan te sterk vergroot is kan dit veroorzaakt worden
door hyperplasie (een toename van het aantal cellen) en door hypertrofie (een vergroting van het celvolume).
Er moet een evenwicht bestaan tussen groeibevorderende en groeiremmende factoren. Deze factoren kunnen door de cel zelf worden
geproduceerd (intracellulaire sturing) of ze worden van buitenaf toegevoegd (extracellulaire sturing). De intracellulaire sturing vindt
plaats via een complex van eiwitten die samen de celcyclus regelen.
Genen die groei en deling stimuleren worden proto-oncogenen genoemd. Deze naamgeving berust op een mogelijke samenhang met
het ontstaan van kanker. Een tumorverwekkend gen wordt een oncogen genoemd (zie hoofdstuk 13). Tumorsuppressorgenen zijn dan
ook genen die groei en deling onderdrukken, afremmen. De extracellulaire sturing vindt plaats door groeistimulerende eiwitten
(groeifactoren) die zich hechten aan het celmembraan en van daaruit
de vorming van bepaalde activatoren tot stand brengen. Groeifactoren
kunnen zowel op lokaal niveau door buurcellen worden gemaakt als
ook afkomstig zijn van andere organen en via het bloed worden
verspreid (hormonen). Dit is bijvoorbeeld belangrijk bij het vormen
van nieuwe weefsels bij genezing na een verwonding. Wanneer de
1
Cellen en weefsels
cellen weer voldoende verbinding met elkaar maken neemt de productie van groeifactoren af.
1.8.3
veroudering
De veroudering berust niet op een mechanisme. Het is een gevolg van
een geleidelijke verandering in de stofwisseling op moleculair en
cellulair niveau. Bij zich delende cellen (bijv. huid of beenmerg) is
apoptose (geprogrammeerde celdood) een normaal verschijnsel.
Door zich te delen voorkomen deze cellen dat zij ouder worden. Het
maximaal aantal delingen per cel (de hayflicklimiet genoemd) is echter
beperkt (gemiddeld ongeveer vijftig). Onder normale omstandigheden speelt dit echter nauwelijks een rol. Zo is de maximumleeftijd die
met deze limiet voor de mens kan worden berekend 122 jaar. Men
denkt dat deze limiet berust op het bij iedere deling korter worden
van de uiteinden van de chromosomen (de telomeren). Het enzym
telomerase dat nodig is voor de verlenging van de telomeren komt
alleen voor in de kiemcellen. Op een gegeven moment zijn de telomeren zo kort geworden dat de celdeling ophoudt.
Langlevende cellen (zenuwcellen, (hart)spiercellen) zijn cellen die,
eenmaal aangelegd, zich niet meer kunnen delen. Hun totaal aantal
staat vast vanaf de kinderleeftijd.
In alle cellen (lang- en kortlevend) vindt voortdurend opbouw en
afbraak plaats van celorganellen en celbestanddelen. Op hoge leeftijd
is de afbraak sterker dan de opbouw met als gevolg verlies van
celfunctie.
1.9
Weefsels
Na de celdeling kunnen cellen zich gaan specialiseren. Dit heet differentiatie. Zoals eerder is vermeld is een weefsel een groep cellen met
dezelfde vorm en functie. Op grond hiervan worden vier typen onderscheiden: dekweefsel, steunweefsel, spierweefsel en zenuwweefsel
(afb. 1.14).
1.9.1
dekweefsel
De cellen in dekweefsel vormen een aaneengesloten laag zonder tussencelstof. Dekweefsel is gelegen aan het lichaamsoppervlak (de
epidermis) en het vormt de binnenbekleding van holle organen, bijvoorbeeld spijsverteringskanaal, luchtwegen en bloedvaten. Dekweefsel begrenst op die manier een holte. Het heeft daarom een
beschermende functie. Om die reden moeten de cellen van het dekweefsel aaneengesloten zijn, zonder tussenruimten. Daarom is er in
dekweefsel geen ruimte voor bloedvaten; het is avasculair. Voeding
vindt plaats vanuit het onderliggende bindweefsel. Omdat dekweefsel
avasculair is kunnen carcinomen in situ (dat wil zeggen: tumoren, die
alleen in het dekweefsel liggen en nog niet door het basaalmembraan
heen gegroeid zijn) niet metastaseren (uitzaaien) naar ander weefsel.
Vanwege het contact met de lichaamsholtes is dekweefsel ook het
enige weefsel dat in contact komt met de inhoud van luchtwegen en
49
Fysiologie en anatomie niveau 5
50
dwarsgestreept (animaal: skeletspieren)
spierweefsel
glad (vegetatief: bijv. wanden maag-darmkanaal
hartspierweefsel
plaveisel
(wand alveoli)
enkellagig
cilindrisch
kubisch
epitheel
(niertubuli)
plaveiselepitheel
meerlagig
dekweefsels
weefsels
met slijmcellen: slijmvlies (bijv. endometrium)
soms met trilharen (tuba, luchtwegen)
soms met microvilli (maag-darmkanaal)
verhoornend (epidermis)
niet-verhoornend (bijv. vagina)
overgangsepitheel
mesotheel
(blaaswand)
(bijv. pleura)
endotheel
straf bindweefsel (pezen, banden, aponeuroses)
(binnenzijde
vaatwand)
elastisch bindweefsel (arteriewanden)
bindweefsels
losmazig bindweefsel (bijv. holten tussen organen)
reticulair bindweefsel (lymfoïde organen)
steunweefsels
vetweefsel (bijv. subcutis)
kraakbeen
hyalien kraakbeen (gewrichtsoppervlakken)
elastisch kraakbeen (bijv. oorschelpen)
beenweefsels
vezelig kraakbeen (bijv. symphysis pubica)
compact (diafyse pijpbeenderen)
zenuwweefsel
spongieus (epifyse pijpbeenderen)
Afbeelding 1.14 Samenvattend schema van weefsels.
maag-darmkanaal. Andere functies van het dekweefsel zijn daarom
ook resorptie en secretie.
Dekweefsel wordt vaak epitheel genoemd, hoewel in feite het epitheel,
naast mesotheel en endotheel, een bepaald type dekweefsel is. Op
grond van de vorm van de dekweefselcellen zijn er verschillende
soorten dekweefsel benoemd (afb. 1.14, afb. 1.15).
Begrenzing betekent het bedekken van het lichaam zowel van buiten als
van binnen. Van buiten gebeurt dit door het verhoornd epitheel van de
huid. Van binnen gebeurt dat door niet-verhoornd dekweefsel, waarbij men drie vormen onderscheidt:
– epitheel; dit vormt het hoofdbestanddeel van de binnenbekleding
van organen die in contact staan met de buitenwereld, bijvoorbeeld
van de luchtwegen en van het spijsverteringskanaal;
– mesotheel (weivlies); dit bestaat uit één laag plaveiselcellen met een
dun laagje bindweefsel met veel zenuwvezels; het vormt de bekleding van longen (de pleurabladen), buikholte (peritoneum parietale) en buikorganen (peritoneum viscerale of serosa). Een tumor,
die uitgaat van het mesotheel, in het bijzonder van de pleurabladen, wordt een mesothelioom genoemd. Dit is een maligne tumor,
die vrijwel altijd wordt veroorzaakt door het inademen van asbestdeeltjes;