Vakfunctionaris open teelt
Examenstandaard: Growing strong 1
Leeractiviteit: Roulerend practicum, plantenhormonen
Doel
Oriëntatie
De functie van plantenhormonen door practicum eigen maken
In dit onderdeel ga je je bezig houden met de verschillende plantenhormonen.
De functie van de verschillende processen wordt in theorie maar ook in
praktijkopdrachten duidelijk gemaakt.
Deze leeractiviteit hoort bij de Oefensituatie Verzorgen van het gewas.
Tijdens het practicum worden de theorie en praktijk aan elkaar gekoppeld
Uitvoering
Bronnen
Afsluiting
Een uitgevoerde praktijkopdracht, met uitgewerkte grafiek.
Beoordelingscriteria
Bijbehorende
Verzorgen van het gewas
competenties
gewasbescherming
Inleiding
"Planten zijn domme, stomme, en passieve organismen die blijven wachten totdat ze worden
opgegeten (door dieren), gemeten (door wetenschappers) of geoogst (door boeren)!".
Deze gedachte dat planten organismen zijn zonder communicatie mogelijkheden, onderling of
met hun omgeving, heeft lang de overhand gehad in de wetenschap. Dit is nog steeds de
algemene gedachte van niet wetenschappers. Niets is minder waar: planten hebben juist een
goed ontwikkeld systeem voor het uitwisselen van natuurlijke hulpbronnen en signalen, ze
kunnen "luisteren" en "praten" met elkaar, maar de manier waarop ze communiceren is
fundamenteel anders dan dat van dieren en mensen.
Het begrip hormoon is in eerste instantie gebruikt in de dierkunde, het heeft daar echter een
andere betekenis dan in de plantkunde, wat blijkt als de werking wordt vergeleken.
Hormonen in zoogdieren:
Hormonen in planten:
Gelokaliseerde productie
Productie vindt plaats in vele organen.
Worden getransporteerd door het bloed naar Transport is geen absolute eis voor de
plaats van actie
werking.
Reguleren de actie door middel van de
Effect van hormonen afhankelijk van
concentratie in het bloed
concentratie en gevoeligheid.
Hormonen bij planten
Deze tekst gaat over hormonen in planten. Welke hormonen zijn er bekend en wat doen ze? Er
worden tot nu toe vijf verschillende groepen hormonen onderscheiden. De hormoonproductie
vindt plaats in “gewone” plantencellen en niet in speciale organen, zoals bij dieren.
Een definitie van plantenhormonen is: “een door de plant zelf gemaakte organische stof, die in
zeer kleine concentraties, de groei en ontwikkeling positief of negatief beïnvloedt”.
Er wordt aangenomen dat een plantenhormoon zich bindt aan een receptor (vaak een eiwit) en
dat de specifieke combinatie van receptor en hormoon de uiteindelijke werking bepaalt.
Plantenhormonen hebben twee soorten werkingen: een snel effect en een langzaam effect. Het
snelle effect is vaak al binnen een half uur zichtbaar. Een voorbeeld hiervan is het effect van
abscisinezuur op het sluiten van de huidmondjes. Het langzame effect is pas na een dag tot
vele weken zichtbaar. Hiertoe behoren de meeste groei- en ontwikkelingsprocessen.
Algemeen geldend voor planten:
Hormonen zijn door de plant geproduceerde stoffen die in lage concentraties regulerend werken
op de fysiologie van de plant.
Hormonen en hun effecten:
Auxine
Zaadkieming
Gibberelline
Cytokinine
Abscisine
Ethyleen
+
+
-
+
-
-
Strekkingsgroei
+
+
Celdeling
+
+
+
Apicale dominantie
+
+
-
Xyleem differentiatie
+
-
Floeem differentiatie
+
Spruitvorming
+
Wortelvorming
+
Bloemvorming
+
-
-
+
+
+
+
Geslachtsbepaling
M
V
+
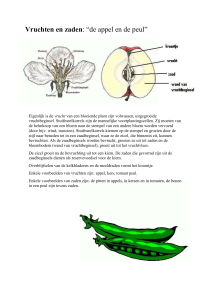
Vruchtontwikkeling
+
+
+
Vruchtval, rijping
-
+
+
Bladval
+
+
+
Veroudering
-
+
+
-
-
+
Net als bij dieren hebben planten hormonen nodig om bepaalde levensprocessen te
beïnvloeden. Er zijn tot nu toe vijf groepen plantenhormonen bekend: auxinen, gibberellinen,
cytokininen, abscisinezuur en ethyleen. Elk hormoon heeft zijn eigen, specifieke bouw en een
specifiek effect op de plant. Een verhoging van het hormoongehalte zal, tot een zeker
maximum, ook een verhoging van het effect tot gevolg hebben.Na de hormoonbinding moet er
een signaal worden doorgegeven van de hormoonreceptor naar de plaats waar het fysiologisch
effect plaatsvindt. Daar kan dan de reeds bestaande eiwitactiviteit beïnvloed worden of een
geheel nieuwe activiteit opgezet worden via DNA-transcriptie. Het effect moet tijdelijk zijn. De
concentratie van een hormoon wordt gereguleerd door een combinatie van synthese, afbraak
en transport. Hormonen kunnen effect hebben op elkaars functioneren. Soms werken ze
samen, soms werken ze elkaar tegen.
Auxine
Al in 1926 toonde Frits Went aan dat er in de top van haver een
stof zit die groei stimuleert. Later werd deze stof geïdentificeerd
als auxine . Het bekendste auxine is indolyl-3-azijnzuur, afgekort
IAA. De belangrijkste syntheseplaatsen zijn: het topmeristeem
van de stengel, vruchtbeginsel, jonge zaden en jonge bladeren.
Vanaf daar wordt het door de plant getransporteerd (derde tekstblok). Auxine is van invloed op
de celwand en bevordert zowel strekkingsgroei die mogelijk is door plastische rekbaarheid als
ook strekkingsgroei door aanmaak van nieuw wandmateriaal.In tegenstelling tot andere
hormonen, is er voor auxine een optimale concentratie , waarboven een verhoging juist een
remmend effect heeft. Het remmende effect van deze verhoging wordt vaak ten dele
toegeschreven aan het vormen van ethyleen waarmee het gepaard
gaat. Naast de natuurlijke auxine zijn er ook vele synthetische
auxines, waarvan 2,4-D de bekendste is. Ze verschillen qua vorm
heel erg met de natuurlijke vorm en danken hun functionaliteit aan
de overeenkomst van een klein deel van de moleculen dat in de
receptor past waardoor de reactie op gang gebracht wordt. Afbraak
kan plaatsvinden onder invloed van licht (foto-oxidatie van auxine)
of onder invloed van enzymen (auxineoxidasen). Auxine heeft
verder een stimulerend effect op de vruchtvorming en vruchtgroei en een remmend effect op de
celdeling in okselknoppen, de zogenaamde apicale dominantie .
Auxine wordt ook gebruikt als herbicide. De werking ligt dan in het toedienen van een overdosis
van synthetische auxines, die veel stabieler zijn dan de natuurlijke waardoor er eerder een
overdosis bereikt wordt.
Auxine scheidt wortel en tak
Hoe het komt dat bij planten de stengel bovenaan zit en de wortels aan de onderkant? Duitse
en Leidse biologen hebben voor het eerst een verklaring gegeven. Met dit belangrijke inzicht in
de basale ontwikkeling van de plant haalden ze de Nature van deze week.
Hester avvn Santen
Voor de vroege scheiding in wortel en stengel blijkt het bekende plantenhormoon auxine
verantwoordelijk. Het onderzoek werd mogelijk doordat plantenembryos sinds kort in de
petrischaal kunnen groeien en door nieuwe methodes die zichtbaar maken waar het hormoon
actief is. Naast de rol van auxine helderden de biologen ook op hoe verschillende genen het
auxine in het vroege embryo de goede kant op sturen.
Al na de eerste celdeling konden de onderzoekers verschillen aanwijzen tussen het
bovengrondse deel van de plant en de rest: in de bovenste cel zit meer auxine. Gedurende de
eerste vijf celdelingen blijft dat zo. Het gen dat die vroege auxinestroom regelt blijkt het nieuw
ontdekte PIN7 te zijn. Na vijf delingen gaan andere PIN-genen aan de slag zodat de
hormoonvloed rechtsomkeert maakt en de ontwikkeling van de wortels begint.
Het feit of stengels al of niet vertakken, wordt in grote mate bepaald door de eindknop
(stengeltop). Deze produceert een plantenhormoon (auxine) dat ervoor zorgt dat de zijknoppen,
die in de bladoksels staan, niet uitlopen (d.i. apicale dominantie). Nochtans is er ook een andere
groep van plantenhormonen belangrijk, want als men cytokininen aan geremde zijknoppen
toedient, gaan deze zijknoppen toch uitlopen.
Voordat zijknoppen kunnen uitlopen tot zijstengels moeten er celdelingen optreden. Hierbij zijn
mogelijk componenten zoals polyaminen betrokken.
Het laboratorium voor plantenfysiologie probeert deze complexe verschijnselen te ontrafelen.
Het werk gebeurt o.a. met tabaksplanten waar een gen (ipt) verantwoordelijk voor de cytokininesynthese (een plantenhormoon) extra werd ingebouwd. Bij deze planten gaat ontbladeren van
de stengel ook gepaard met het opheffen van apicale dominantie. Bij tabaksplanten werden
veranderingen in polyaminen gevonden, tezamen met zeer grote stijgingen van tyramine en
fenethylamine. De functie van deze veranderingen is nog niet bekend.
Ook bij bonen wordt apicale dominantie bestudeerd. Bij afsnijden van de apex (decapitatie)
lopen de zijknoppen uit. Hierdoor gaat ook het gehalte aan putrescine en spermidine stijgen.
Jiri Friml e.a.: ‘Efflux-dependent auxin gradients establish the apical-basal axis of
Arabidopsis‘. Nature, 12 november
Gibberelline
Er zijn vele verschillende natuurlijke gibberelines ontdekt en deze zijn chemisch gezien in
hoge mate gelijk aan elkaar . De meest actieve GA is GA1. Veel andere gibberellines danken
hun activiteit vooral aan hun mogelijkheid omgezet te worden tot GA1. Ook gibberelline is een
groeibevorderend plantenhormoon (celdeling en celstrekking) en vooral essentieel bij de
stengelgroei. Veel planten die met gibberelline behandeld worden vertonen abnormale
lengtegroei. Doordat bekend is dat planten die een genetisch defect hebben in de
gibberelline-aanmaak dwerggroei vertonen, kan geconcludeerd worden dat alle planten in
hun eigen gibberelline-aanmaak voorzien. Gibberelline wordt vooral gevormd in de jonge
bladeren, de zaden en in de jonge wortel (top). Het transport vindt plaats via het
vaatbundelweefsel vanuit de bladeren naar de jonge groeiende delen. Het speelt een
belangrijke rol in het doorschieten van tweejarige planten. Ook het uitlopen van de knoppen
in het voorjaar is afhankelijk van GA. In sommige planten stimuleert GA de vruchtzetting.
Zaden hebben voor hun kieming een koudeperiode (zoals de winter) of een lichtbehandeling
(zoals langer wordende dagen) nodig. GA kan deze behandelingen vaak vervangen. Bij
zaden heeft GA invloed op de mobilisatie van reservestoffen bij de kieming. Hierdoor komen
de reservestoffen van de ontwikkelende kiem ter beschikking.
Cytokininen
Cytokininen zijn in eerste plaats bekend als
celdelinghormonen die samenwerken met auxine. Aan deze
functie dankt het ook zijn naam; cytokinese is celdeling.
Door
deze eigenschap wordt het vaak toegepast in stekpoeder.
Cytokininen worden synthetisch nagemaakt , maar de
effectiviteit hiervan is vaak veel lager dan van de natuurlijke.
Productie van cytokinine vindt plaats in jonge organen en in de wortelpuntjes. Van hieruit
wordt het passief via de houtvaten getransporteerd.Een combinatie van voedingsstoffen,
auxine en cytokinine kunnen er voor zorgen dat ook gedifferentieerde cellen weer in staat zijn
om te gaan delen. Er kunnen zelfs ongedifferentieerde cellen ontstaan die blijven delen:
calluskweek. In combinatie met auxine bevorderen cytokininen de aanleg van wortels en
spruiten. De verhouding tussen auxine/cytokinine bepaalt welke onderdelen er gevormd
worden. Cytokinine is ook effectief tegen het vergelen van bladeren. In de sierplantenteelt
geeft dit een bruikbare oplossing van een commercieel probleem.
Abscisinezuur
In tegenstelling tot auxine, gibberelline en cytokinine, heeft
abscisinezuur (ABA) vaak een remmende werking op een
proces. ABA wordt voornamelijk gesynthetiseerd in de
chloroplasten van oudere bladeren, maar ook in worteltoppen,
vruchten en zaden, alwaar het de kieming remt. ABA kan de
aanmaak van ethyleen bevorderen. Het werkt voornamelijk als antagonist van de eerder
beschreven hormonen. De kiemrust van zaden, bedoeld om te voorkomen dat zaden gaan
ontkiemen in een ongunstige periode zoals de winter, is een gevolg van de aanwezigheid van
ABA. Het effect op de celdeling ligt in het afremmen van de eiwit- en RNA-synthese. Bij het
werkingsmechanisme van het sluiten van de huidmondjes ligt het anders. Het effect van ABA
bij het stimuleren van het sluiten van de huidmondjes is al binnen acht minuten zichtbaar,
waardoor een rol voor eiwit- of RNA-synthese is uitgesloten.
Ethyleen (H2C=CH2)
Ethyleen kan in alle organen gesynthetiseerd worden en is met name in bloemen en vruchten
een katalysator voor de aanmaak van nog meer ethyleen. Ook hoge concentraties van auxine
stimuleren de aanmaak van ethyleen. Het gevormde ethyleen wordt als gas getransporteerd
via de intercellulaire holtes. Het kan snel worden afgegeven en zo van plant naar
plant overgaan om ook daar een effect te induceren zoals bijv. vruchtrijping. De essentiële
rol hierbij is aangetoond m.b.v.transgene planten die geen ethyleen meer produceerden.
Deze planten vormden wel vruchten, maar deze werden niet rijp. Bij het rijp worden van
vruchten verandert het vruchtweefsel, er ontstaan kleur- en geurstoffen en verzoeting door de
vorming van suikers. Ethyleen is van invloed op de DNA-transcriptie, waardoor
celwandafbrekende enzymen worden geproduceerd zodat de vruchten zacht worden. Fruit
rijpt ook als het niet meer vastzit aan de plant. Hiervan wordt gebruik gemaakt bij het bewaren
en vervoeren ervan. Buiten vruchtrijping heeft ethyleen nog andere effecten op
groeiprocessen in de plant. Het bevordert een aantal verouderingsprocessen zoals
bloemverwelking en bladval. Daarnaast is ethyleen een stresshormoon.
Proeven met groeihormonen
Inleiding
Bij de commerciele plantenteelt wordt vaak gebruik gemaakt van verschillende soorten
kunstmatige groeistoffen. Door toevoeging van deze hormonen wordt de groei versneld en
daarme de productie verhoogd. In onderstaande proefjes ga je het effect van 2 van deze stoffen
bestuderen.
De invloed auxine op de lengtegroei van tuinkers
Benodigdheden
- auxine (= 2,4D = 2,4
dichloorfenoxyazijnzuur, molecuulgewicht 221)
- aquadest.
- 5 kleine erlenmeyers
- Finn-pipet + tip
- 6 petrischalen
- filtreerpapier
- tuinkerszaadjes
- lineaal
Tuinkers
(Lapidium sativum)
Werkwijze
Bereid een oplossingsreeks van 2,4 D van 5 verschillende concentraties: 10-15M, 10-12M,
10-10M, 10-5M en 10-3M. De 6e petrischaal wordt ingezet als controle, dus zonder
groeihormoon.
Voeg 10 ml van elke concentratie toe aan met filtreerpapier beklede petrischalen waarin
steeds 10 tuinkerszaden gelegd zijn.
Laat de zaden in het donker kiemen en bepaal na 1 week per schaaltje de gemiddelde
lengte van de stengel en van de wortel, van alle gekiemde zaden.
Zet de gegevens uit in een grafiek. X-as: concentratie 2,4 D. Negatieve Y-as: lengte van
de wortels. Positieve Y-as: lengte van de stengels.
Vragen
Welke concentratie 2,4 D beinvloedt de lengtegroei van de stengels en de wortels
optimaal?
Waarom is er ook groei zonder hormoon toevoeging?
B. De invloed van gibberelline op de groei van sla
Benodigdheden
- slazaad
petrischalen
gibberelline (GA)
kleine erlenmeyers
filtreerpapier
Finn-pipet + tip
thermometer
aquadest
lineaal
-6
-5
-
Sla
(Lactuca sativa)
Werkwijze
Zaai slazaad uit in met vochtig filtreerpapier beklede petrischalen en laat de zaden
kiemen bij ca 25 C, zet de schaaltjes in het licht. De zaailingen zijn bruikbaar als de
worteltjes 5-6 mm lang zijn.
Maak een oplossingsreeks uitgaande van 100 mg/I GA, zodat concentraties verkregen
worden van 10mg/l; 1mg/l; 0,1mg/1 en 0,01mg/l.
Bekleed 6 petrischalen met filtreerpapier, voeg 5ml van elke oplossing toe. Noteer per
schaaltje welke concentratie wordt toegevoegd. De 6e wordt uitgezet als controle.
Leg in elke schaal 20 kiemende slazaden. Laat de schalen bij zwak licht staan,
gedurende 4-5 dagen. Met water bijvullen als het papier droog dreigt te worden.
Meet de lengte van alle hypocotylen. Zet de gevonden (gemiddelde) waarden uit tegen
de logarithme van de GA-concentratie.
Vraag: Is de groei evenredig met de hormoonconcentratie?
Ethyleen
Inleiding.
Snijbloemen verwelken sneller als er fruit in de buurt is, een weetje uit grootmoeders tijd dat
erom vraagt nader onderzocht te worden. Als bloemen in het experiment worden anjers gebruikt
daar deze extra gevoelig zijn. Voor het fruit wordt gekozen voor de appel. Door deze keuzes is
getracht de normale situatie na te bootsen.
Theorie
Ethyleen werd bij toeval ontdekt door citroenkwekers. Citroenen worden groen geoogst. Om ze
hun gele kleur te laten krijgen werden de citroenen verwarmt met kerosinekachels in de
veronderstelling dat warmte de citroenen geel kleurt. Bij een andere manier van verwarmen
bleven de citroenen geel. Nader onderzoek wees uit dat de citroenen niet geel kleurde o.i.v.
warmte, maar o.i.v. een kleine hoeveelheid ethyleen die vrijkwam bij het verbranden van
kerosine. Ethyleen is een gasvormig plantenhormoon. Ethyleen is een gas dat vrijkomt uit
rijpend fruit en het is verantwoordelijk voor het verwelken van snijbloemen. De molecuulformule
voor het gas ethyleen is C2H4. Andere plantenhormonen zijn cytokininen, gibberrellinen en
auxine. Waar ethyleen een negatief effect heeft op het vaasleven van snijbloemen hebben de
andere hormonen hier juist een positief effect op. Voor de vorming van cytokinine is suiker
nodig. Daardoor verlengt het toevoegen van suiker aan het water de levensduur van
snijbloemen.
De anjer is erg gevoelig voor ethyleen. De gevoeligheid van een snijbloem lijkt af te hangen van
het aantal receptoren voor ethyleen dat de soort heeft. Meer receptoren leidt tot een hogere
gevoeligheid. Bij de afgesneden anjer wordt door ethyleen dat van buiten de bloem
binnendringt, de eigen ethyleenproductie sterk gestimuleerd. Deze snel oplopende
ethyleenproductie zorgt ervoor dat de anjer verwelkt. Ethyleen versnelt het verouderingsproces.
Materiaal en methode
De bloemen en de appels moeten in een kleine, luchtdicht afgesloten ruimte bij elkaar worden
geplaatst. Uit praktische en financiële overwegingen is er gekozen de opstellingen in grote
doorzichtige plastic zakken te plaatsen. De gekozen bloemensoorten zijn de anjer (zeer
gevoelig voor ethyleen) en de chrysant (vrij ongevoelig voor ethyleen. Als appelsoort is er
gekozen voor de Elstar (zonder een duidelijke reden). De appels werden in een weckpot
geplaatst om daarin ethyleen te 'kweken'. Een weckpot is luchtdicht af te sluiten en door de
rubberring kan met een injectienaald gas worden afgenomen.
Om het gas te onderzoeken is gebruik gemaakt van een gaschromatograaf. Voordat het gas in
de gaschromatograaf is gespoten werd het door een zeolietbuis geleid om het gas te drogen.
Zeolietkristallen absorberen waterdamp. De gaschromatograaf is via een signaalversterker
gekoppeld aan het meetpaneel en met het programma IP-coach zijn m.b.v. de computer de
metingen uitgevoerd. Met ethyleengas is een ijkgrafiek gemaakt, deze ijkgrafiek is vergeleken
met het in de weckpot 'gekweekte' gas. De bloemenopstelling:
5 oranje anjers en 10 elstar appels in één plastic zak
5 oranje anjers in één plastic zak (blanco)
5 witte anjers in één plastic zak; 25 ml ethyleengas geïnjecteerd
5 witte anjers in één plastic zak (blanco)
5 witte chrysanten en 10 elstar appels in één plastic zak
5 witte chrysanten in één plastic zak (blanco)
Figuur 1: De opstellingen.
Resultaten
Grafiek 1 is de ijklijn van het gas ethyleen, grafiek 2 is de lijn die de gaschromatograaf geeft bij
inspuiting van het gas uit de weckpot dat is geproduceerd door de appels.
Grafiek 1: ethyleen
Grafiek 2: appelgas
De grafieken geven op hetzelfde punt een ongeveer evengrote piek te zien. Hieruit kan worden
geconcludeerd dat het gas dat de appels produceren ethyleen is.
Na 10 dagen in de opstelling worden de resultaten op het oog vastgesteld:
5 oranje anjers en 10 elstar appels in één plastic zak:
bloemen zijn verwelkt
5 oranje anjers in één plastic zak (blanco):
bloemen zijn niet verwelkt
5 witte anjers in één plastic zak; 25 ml ethyleengas geïnjecteerd: bloemen zijn niet verwelkt
5 witte anjers in één plastic zak (blanco):
bloemen zijn niet verwelkt
5 witte chrysanten en 10 elstar appels in één plastic zak:
bloemen zijn niet verwelkt
5 witte chrysanten in één plastic zak (blanco):
bloemen zijn niet verwelkt
Discussie
De oranje anjers die samen met de appels in één plastic zak zaten zijn verwelkt, de oranje
anjers zonder de appels zijn niet verwelkt. Hieruit kan worden geconcludeerd dat het door de
appels geproduceerde ethyleengas de anjers doet verwelken. Opmerkelijk is dat de witte anjers
in de zak waarin ethyleengas is geïnjecteerd niet zijn verwelkt. Dit is mogelijk te wijten aan het
feit dat ethyleen vrij snel de plastic zak heeft kunnen verlaten (mogelijk door diffusie of door het
niet perfect afgesloten zijn van de zak). Bij de proef met de appels speelde dit een kleinere rol
omdat de appels voor een voortdurende aanvoer van ethyleengas verantwoordelijk waren. Uit
de vergelijking tussen de oranje anjers met appels in de zak en de witte chrysanten met appels
in de zak kan worden geconcludeerd dat de anjers gevoeliger voor ethyleengas zijn dan de
chrysanten.
Er kan worden vastgesteld dat rijpend fruit een nadelige invloed heeft op het vaasleven van
anjers. Het weetje uit grootmoeders tijd is hiermee bewezen. Echter één ding heeft grootmoeder
over het hoofd gezien, het geldt niet voor alle snijbloemen in even sterke mate. Zo zijn
bijvoorbeeld chrysanten veel minder gevoelig. Maar ja, wat wist grootmoeder nou van
ethyleenproductie? Daar heeft zij haar meer ontwikkelde achter-achter-achter-achter
kleindochters voor nodig.
EXTRA INFO BESCHIKBAAR indien nodig !!!
Auxinen
Belangrijke productie plaatsen van auxinen zijn: Het topmeristeem van de stengel,
vruchtbeginsels jonge bladeren en jonge zaden.
Auxine wordt door de bladgroencellen (parenchymcellen) die in contact staan met de
vaatbundels, van jonge blaadjes naar beneden getransporteerd. Dit transport verloopt langzaam
(1 cm/uur), in de stengel naar de basis vd plant en in de wortel naar de punt van de wortel. Het
transport van auxine kost energie.
Inactivatie in de plant vindt plaats door conjugatie met suikers of oxidatie.Auxine bevordert de
celstrekking, cambiumactiviteit, vruchtvorming en vruchtgroei, en speelt een rol in de knoprust.
Verwijdering van jonge bladeren en knoppen remt de vorming van nieuwe wortels. Wordt
vervolgens auxine toegediend, dan worden de nieuwe wortels alsnog gevormd. Auxine
bevordert de vorming van wortels aan stengels (=adventiefwortels). Auxinen worden daarom in
allerlei formuleringen van stekpoeder gebruikt. In hoge concentraties veroorzaakt IAA de
productie van ethyleen. De werking van auxine wordt beïnvloed door de aanwezigheid van
cytokinine.
Gibberelline
Gibberelline wordt vooral in de jonge bladeren geproduceerd, net zoals de auxinen. Wortels
produceren eveneens gibberelline. Transport vanuit de wortel naar de top vindt plaats via het
xyleem, vanuit de bladeren via het floeem naar de jonge groeiende delen.
Wanneer gibberelline aan de wortel oplossing wordt toegevoegd, wordt de vorming van
zijwortels onderdrukt. In het algemeen werken gibberellinen groeiversnellend en ze verlengen
de strekkingsgroei van de intacte plant. Gibberelline worden onder andere gebruikt om
onbevruchte peren en druiven volledig te laten ontwikkelen. Toevoegen van gibberelline kan
een koude of licht behandeling voor de kieming van zaden vervangen. Bij de groei van de wortel
speelt gibberelline geen rol.
Inactivatie van gibberelline vindt plaats door conjugatie aan suikers of door omzetting in een
minder actieve gibberelline.
Cytokinine
Cytokinine concentratie is het hoogst in jonge organen (zaden, vruchten en bladen) en in de
wortelpuntjes. Op deze plaatsen en in de stengel wordt de cytokinine geproduceerd. Transport
vindt vooral plaats via het xyleem.
Cytokinine activeert de stofwisseling, bevordert de celdeling, doorbreekt rust in knoppen en
zaden. In callus weefsel ontstaan knoppen, stengels en bladen, wanneer de verhouding
Cytokinine/auxine hoog is. Is deze verhouding laag dan ontstaan complete planten. Is er een
overmaat aan auxine dan ontstaan wortels. Cytokinine vertraagt de veroudering, zij beschermt
de membranen tegen oxidatie door zuurstof radicalen. Hoge cytokinine concentraties in een
orgaan of weefsel brengen een transport opgang van assimilaten naar dat weefsel of orgaan.
De productie van cytokinine in de wortel zorgt er voor dat de suikers en andere voedingsstoffen
naar de wortel getransporteerd worden.
Inactivatie van cytokinine vindt plaats door conjugatie met een suiker of door oxidatie. De
glucosides van cytokinine zouden een opslag en transport vorm kunnen zijn.
Ethyleen
Ethyleen wordt door alle organen gevormd. In de grond is de ethyleen niet alleen van de wortels
afkomstig. Ook bacteria en schimmels maken ethyleen, in de grond kunnen deze organismen
dus een hormonale invloed op de plant uitoefenen.
Transport van ethyleen vindt plaats via de intercellulaire ruimtes. Inactivering vindt plaats door
oxidatie, tevens kan ethyleen als gas aan de atmosfeer worden afgegeven. Ethyleen kan dus
vrij eenvoudig verdwijnen. In het geval dat dit onmogelijk is, kunnen afwijkingen ontstaan door
ophoping van ethyleen. Dit is bijvoorbeeld het geval wanneer de wortels langdurig onder water
staan. In deze omstandigheden is de productie van ethyleen weliswaar verminderd, maar de
afvoer van ethyleen staat vrijwel geheel stil. De verschijnselen die dan optreden zijn:
bladchlorose, versterkte strekking van de stengel, gecombineerd met een verdikte stengel,
slaphangen, naar de stengel buigen van de bladen en eventueel bladval, afnemende
wortelstrekking, vaak gecombineerd met de vorming van adventief wortels, en een toegenomen
gevoeligheid voor ziekteverwekkers.
De verhoogde ethyleen concentratie in waterverzadigde grond kan in de schorscellen de
vorming van cellulase opwekken. Hierdoor lossen de celwanden op. De celinhoud verdwijnt
eveneens, zodat een holte overblijft. Het aerenchym weefsel ontstaat op deze wijze.
De ethyleenovermaat verschijnselen bovengronds zijn een indirect effect van het ondergrondse
zuurstofgebrek. De voorloper van ethyleen ACC heeft zuurstof nodig om ethyleen te vormen.
Deze stof wordt via het xyleem naar boven getransporteerd en daar in de zuurstofrijke. Bij rozen
is deze situatie nog niet gerapporteerd.
Onder normale omstandigheden remt ethyleen de strekking en verdikt de wortel en de stengel.
Dit is goed zichtbaar tijdens de kieming.
Abscisinezuur
Abscisinezuur remt de celdeling en de celstrekking en remt het uitlopen van rustende knoppen
en zaden, het kan als antagonist van de gibberelline, auxine en cytokinine beschouwd worden.
Het is een remstof van de groeiprocessen.
Abscisinezuur wordt gevormd door de afbraak van carotenoïden. Belangrijke productie plaatsen
zijn de bladgroenkorrels in verouderend blad. Het wordt eveneens geproduceerd in de
worteltoppen, vruchten en zaden. Transport vindt plaats via het xyleem en floeem. De
belangrijkste transportweg is vanaf het blad via het floeem naar de sinks.
Abscisinezuur wordt geïnactiveerd door oxidatie en door de conjugatie met suikers.
Is de aanvoer van Abscisinezuur in de groeipunten groot genoeg dan stopt de celdeling in de
groeipunten van wortel en spruit, terwijl op de ogen knopschubben ontstaan. De plant gaat in
rust.
Transport in planten
Planten groeien mede onder invloed van het groeihormoon auxine. Auxine bevordert de
celstrekking en wordt onder andere in de stengeltop van een plant geproduceerd. In een
onderzoek naar het transport van auxine worden vier experimenten gedaan.
In experiment 1 wordt een stukje stengel van een plant in drie delen verdeeld: de stengeltop,
het middelste deel en het onderste deel. Vervolgens worden de delen weer in dezelfde positie
op elkaar gezet. Hierna groeit de stengel door (zie de afbeelding hieronder).
In experiment 2 wordt een stukje stengel op dezelfde manier in drie delen verdeeld. Vervolgens
wordt het middelste deel omgekeerd teruggeplaatst. Hierna groeit de stengel niet door (zie de
afbeelding hieronder).
auxine
Op grond van de resultaten van de experimenten 1 en 2 worden twee vervolgonderzoeken
opgezet, de experimenten 3 en 4. In deze experimenten wordt het middelste deel van een
overeenkomstig stukje stengel tussen twee blokjes agar geplaatst. Agar is een stof die auxine
doorlaat. Aan het bovenste blokje is auxine toegevoegd, aan het onderste blokje niet (zie de
afbeelding hieronder).
In experiment 3 wordt het stengeldeel rechtop geplaatst. Na enkele uren blijkt auxine in het
onderste blokje agar te zijn gekomen.
In experiment 4 wordt het stengeldeel omgekeerd geplaatst. Nu komt geen auxine in het
onderste blokje terecht (zie de afbeelding hieronder).
auxine
Formuleer een onderzoeksvraag die met de resultaten van de experimenten 3 en 4 kan worden
beantwoord.
Drie leerlingen trekken op grond van de resultaten van de experimenten 1, 2, 3 en 4 de
volgende conclusies:
leerling 1: het transport van auxine vindt plaats door de bastvaten;
leerling 2: het transport van auxine vindt plaats van de top van de stengel naar het onderste
deel van de stengel;
leerling 3: het transport van auxine vindt plaats van de top van de stengel naar het onderste
deel van de stengel door diffusie.
Welke van deze leerlingen trekt een juiste conclusie uit de experimenten 1, 2, 3 en 4?
A: geen van deze leerlingen
B: alleen leerling 3
C: alleen de leerlingen 2 en 3
D: alleen leerling 2
E: alleen leerling 1
F: alleen de leerlingen 1 en 3
http://www.nice-info.be
http://www.rhizopon.
com/historie1.html#
1
http://www.digischo
ol.nl/bioplek/animati
es/cel/celstrekking.h
tml
http://www.biologie.
uni-hamburg.de/bonline/e31/31d.htm
http://www.vcbio.sci
ence.ru.nl/fesem/applets/vein/
http://www.plant.dlo.nl/news/archive/pb98-9.html