lipiden")
Celbiologie 7 september 2011
dr. Van der Klei
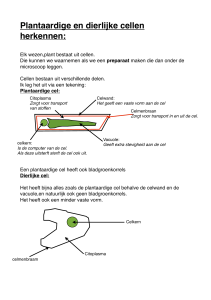
(Fosfo)lipiden: Hydrofiele kop, met
hydrofobe (vet)zuurketens. Dit zijn de
bouwstenen van verschillende membranen
in een cel. Zo is bijvoorbeeld het
celmembraan opgebouwd uit deze
fosfolipiden (de bi-layer. De celkern (en ook
andere organellen) heeft 2 bi-layers, de
inner en de outer layer. Deze lipiden worden
gemaakt door enzymen! Onthoud wel, dat
een membraan, uit lipiden is opgebouwd en
een celwand uit suikers.
Eiwitten: Opgebouwd uit aminozuren.
Eiwitten kunnen verschillende functies
hebben. Ze kunnen bijvoorbeeld werkzaam
zijn als enzym, een receptor of als een
structureel eiwit dienen (bijvoorbeeld poriën in een membraan). Een werkend eiwit, bestaat eigenlijk
uit verschillende, kleinere eiwitten (quatenaire structuur). De opvouwvolgorde van een eiwit ligt al
vast in het DNA. Hierdoor liggen de waterstofbruggen op een dusdanige volgorde, dat de
opvouwvolgorde altijd hetzelfde zal zijn.
Omdat een celmembraan zo bewegelijk is, kan het voorkomen dat bepaalde eiwitten diffuseren over
het oppervlak van het celmembraan. Om dit te voorkomen (bijvoorbeeld bij een darmcel, waar je wil
dat bepaalde soorten eiwitten, juist aan de
laterale of de basale kant van de cel blijven)
heeft moeder natuur de zogenaamde “tight
junctions” bedacht. Door deze junctions (wat
in feite ook gewoon eiwitten zijn), is het voor
eiwitten onmogelijk om over de gehele cel te
diffuseren.
Eu/prokaryoot: Eukaryoten zijn alle
organismen waarvan de cellen een celkern en
een intern membraan hebben. In de
taxonomie worden ze beschouwd als één van
de drie domeinen waarin het leven verdeeld
kan worden. De andere twee domeinen, de Archaea en Bacteria, worden samen prokaryoten
genoemd en hebben een "simpeler" celbouw waarin het DNA niet omsloten wordt door een celkern
maar vrij in het cytoplasma ligt.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
1
Op DNAniveau lijken eukaryoten relatief veel
op elkaar. Een eencellige gistcel lijkt op dat
niveau bijvoorbeeld meer op een mens dan op
een bacterie. De grootte van een cel/organisme
zegt dus niks over de complexiteit van het DNA.
Hierdoor wordt gist veel gebruikt in het
onderzoek naar kanker. Vooral de mens en een
witte muis lijken erg op elkaar. In de afbeelding hiernaast hebben de muis en de mens dezelfde
mutatie in het DNA, waardoor zij beiden een witte vlek op het voorhoofd hebben. Het DNA op de
plek van de mutatie zorgde dus blijkbaar voor het pigment op die plek.
Perixosomen: Een peroxisoom is een celorganel in een eukaryote cel. Een enkel membraan sluit hem
af van het cytosol (interne vloeistof van een cel).Peroxisomen zijn in staat waterstofperoxide (H2O2)
te vormen door waterstof bij zuurstof te plaatsen. Dit kan verschillende functies hebben.
Peroxisomen staan in voor de synthese van bepaalde fosfolipden die betrokken zijn voor de
efficiënte geleiding van impulsen in zenuwcellen. Peroxisomen in de lever zijn bijvoorbeeld in staat
giftige stoffen zoals alcohol te detoxificeren. Bij dit proces wordt het waterstof van de gifstoffen bij
zuurstof geplaatst waarmee waterstofperoxide gevormd wordt. Waterstofperoxide is zelf ook giftig,
maar peroxisomen bevatten enzymen om deze stof om te zetten in water en zuurstofgas.
Progeria is een zeldzame, autosomale dominante verouderingsziekte die wordt veroorzaakt door een
mutatie in het LMNA-gen. De klassieke vorm van progeria staat bekend als het Hutchinson-Gilford
Progeria Syndroom (HGPS), genoemd naar de artsen Hutchinson en Gilford die de ziekte voor het
eerst beschreven in 1886 en 1904. De naam
progeria komt uit het Grieks en betekent: "sneller
oud worden". In de afbeelding zie je het verschil
tussen een normale celkern (boven) en een celkern
van een Progeriapatient. Of er nog verder wordt
ingegaan op deze ziekte (of op de dia’s in de
sheetmap) is mij nog onduidelijk.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
2
Celbiologie 12 september 2011
dr. P van Haastert
Cellulaire communicatie
Principe van cel-cel communicatie
Een hormoon koppelt zich aan een receptor op het cel
membraan. De receptor geeft een signaal af aan de
binnenkant van het membraan (via
signaleringseiwitten). De cel kan hier op verschillende
manieren op reageren.
-De cel kan een verandering het zijn metabolisme
ondergaan
-Er kan een verandering in de genexpressie optreden
(de mate waarmee het DNA van een gen gekopieerd
wordt naar messenger RNA (mRNA) of andere RNA's
en mRNA vertaald wordt naar een
aminozuursequentie.)
-Ook kan er een verandering optreden in de vorm of beweging van de cel.
Manieren van signalering
Signaalmoleculen (bijvoorbeeld hormonen of neurotransmitters) kunnen op verschillende manieren
werkzaam zijn. Ten eerste is er de contactafhankelijke overdracht. Hierbij zit het
signaalmolecuul op het membraan van een
cel, die vervolgens het receptormolecuul
op de doel cel kan activeren, waardoor
deze cel één van de bovengenoemde
acties gaat ondernemen. Bij paracrine
signalering reageren cellen op
signaalmoleculen die door een andere cel
zijn uitgescheiden. Bij synaptische
signalering worden de receptoren op het
membraan van de doel cel geactiveerd,
omdat het uiteinde van een zenuwcel
verschillende neurotransmitters uitscheidt.
Bij de laatste vorm van signalering wordt
er gebruikgemaakt van het
bloedvatenstelsel. Hierbij scheidt een cel
signaalmoleculen uit, waar andere cellen
op een andere plaats in het lichaam weer op reageren, deze manier van signalering heeft dus een
veel groter bereik dan de anderen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
3
Endocriene en synaptische signalering
Bij endocriene signalering worden er veel verschillende signaalmoleculen (hormonen) in de
bloedbaan uitgescheiden. Omdat er ook veel targetcellen zijn, zal elk hormoon weer op een
specifieke receptor ‘passen’. Hierdoor zal een hormoon dat werkzaam moet zijn in de lever, ook
alleen in de lever werken, omdat die hormonen een dusdanige vorm hebben dat ze alleen op
levercelreceptoren passen.
Bij synaptische signalering werkt het allemaal weer net even anders. Omdat elk post synaptisch
membraan één bepaald soort neurotransmitter uit kan scheiden, is het belangrijk dat deze precies
uitkomen bij de cellen (en dus de receptoren) waar deze neurotransmitters op passen. Je moet er
dus voor zorgen dat neuron A, uitkomt bij doelcellen A en neuron B bij doelcellen B etc.
Effecten van een signaal molecuul op een cel
Zoals ik al zei ka een cel op verschillende manieren reageren op een extracellulair signaal molecuul.
Dit kan via een relatief snelle en een wat langzamere weg. Via signaaltransductie (=het doorgeven
van signalen binnen de cel) kan het cytoplasma zich anders gaan gedragen, wat weer kan leiden tot
een verandering in het gedrag van de cel. Een wat langzamere weg is de weg van de eiwitsynthese.
Bij deze weg zet de geactiveerde receptor op het membraan de cel aan het werk om een bepaald
eiwit te maken.
Gap Junctions
Dit zijn kanaaltjes in het celmembraan die doorlaatbaar zijn tot
1000Da (d.w.z. geen eiwitten of DNA). Deze Gap Junctions zijn
belangrijk voor o.a. elektrische koppelingen tussen cellen in het hart.
Extracellulaire signalen
Cellen zijn altijd afhankelijk van extracellulaire
signalen. Zoals je in het plaatje hiernaast
kunt zien heeft een cel (in dit geval A,B en C)
nodig om te overleven. Als een cel wil delen
(en dit doet hij niet zodra er genoeg
voedingsstoffen zijn) moet hij eerst
signaalmoleculen D en E ontvangen. Ook dan
moet de cel A,B en C blijven ontvangen. Zodra
de cel A,B en C niet meer ontvangt, zal de cel
doodgaan (apoptose).
Een levercel zal A,B en C van cellen in zijn
omgeving ontvangen. Mocht je nou een long
cel tussen deze levercellen plaatsen, dan zou
deze long cel doodgaan omdat het soort A,B en C afhankelijk is van de soort cel. Een long cel heeft
dus LONG-A,B en C nodig i.p.v. LEVER-A,B en C.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
4
Verschillende reacties op een stof
Cellen kunnen verschillend reageren op een stof (bijvoorbeeld acetylcholine). Zo zorgt acetylcholine
dat een hartspier ontspant, maar dat een skeletspier juist aanspant. In speekselklieren zorgt
acetylcholine ervoor dat de klier speeksel uitscheidt. Omdat acetylcholine een neurotransmitter is,
spreken we dus van synaptische signalering.
Typen signaalmoleculen
In de celbiologie spreken we van 3 verschillende typen signaalmoleculen. Zo hebben we hydrofobe,
hydrofiele en gasvormige signaalmoleculen.
Hydrofobe signaalmoleculen
Omdat sommige signaalmoleculen hydrofoob zijn, kunnen ze gemakkelijk door het celmembraan
heen. Daar kunnen ze koppelen aan nucleaire receptoren (elk hydrofoob signaalmolecuul heeft een
eigen receptor!) Elk van deze receptoren wordt actief zodra deze bindt aan een signaalmolecuul.
Zodra dit gebeurt, zal er DNA transcriptie plaatsvinden en zal er bijvoorbeeld een eiwit gevormd
worden.
Gasvormige signaalmoleculen
In het centrale zenuwstelsel is NO een neurotransmitter, en in het autonome zenuwstelsel is NO een
neurotransmitter die gladde spiercellen ontspant, onder andere in het maagdarmkanaal en in
bloedvaten (vaatverwijding, betere doorbloeding). Omdat NO gas zo reactief is, heeft het een korte
levensduur.
Transductie van hydrofiele signalen
Er zijn grofweg gezegd 3 manieren waarop
hydrofiele signalen een cel tot actie aan
kunnen zetten. Ten eerste zijn er de
zogenaamde ionen kanalen. De grootte van
de opening van deze kanalen bepaalt welke
ionen er wel of niet doorkunnen. Deze ion
poorten gaan pas open wanneer er een signaalmolecuul zich aan een receptoreiwit op het poortje
bindt.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
5
G-eiwit gebonden receptoren
G-proteïnegekoppelde receptoren zijn receptoren die zich in het
celmembraan bevinden, en signalen kunnen ontvangen van de buitenkant
van het membraan, en een respons geven aan de binnenkant van het
membraan. De receptor aan de binnenkant van het membraan activeert een
G-proteïne, die vervolgens een cascade van andere reacties in gang zet en zo
een verandering elders in de cel bewerkstelligt.Het G-Proteïne bestaat uit 2
delen namelijk een α- en βγ-deel. Wanneer het G-proteïne inactief is bindt
het α-deel met GDP. Het βγ-deel zit dan aan het α-deel vast. Als het Gproteïne actief is, is het gebonden met GTP en het α- en βγ-deel zijn van
elkaar gesplitst. Het α-deel en/of βγ-deel kunnen dan reageren met een
effector. Het G-proteïne werkt samen met de G-proteïnegekoppelde receptor
(GPCR).
Zodra het α- en βγ-deel geactiveerd is, zal het alfa deel zich afsplitsen van het βγ-deel, omdat de
ruimtelijke bouw veranderd. Het het α-deel zal zich vervolgens hechten aan het doeleiwit (enzym),
zoals ik hieronder heb weergegeven. In de tekening staat dat het bèta-gamma deel ook nog aan het
alfa deel blijft vastzitten, ik weet niet wat het nou is, elke website zegt wat anders.
Laten we doorgaan. Zodra het αdeel met gebonden GTP (want het
is een geactiveerd α-deeltje) zich
heeft gehecht aan het enzym
(adenylyl cyclase), wordt het enzym
actief. Dit enzym zorgt ervoor dat
ATP wordt omgezet in cAMP
(cyclisch AMP). Dit cAMP hecht aan
een inactief PKA-molecuul. Zodra
dit gebeurd, splitsentwee actieve onderdelen van het PKA-molecuul af (active catalystic subunits, zie
hieronder/naast). Deze twee subunits gaan in endocriene cellen de nucleus in, waar ze met behulp
van ATP, een inactief CREB-molecuul fosforyleren, waarbij een ADP vijkomt. Dit gevormde complex
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
6
zal vervolgens nog hechten aan een CREB-bindingseiwit (CBP). Dit uiteindelijk gevormde complex zal
in staat zijn om DNA transcriptie uit te voeren en dus in staat zijn om eiwitten te produceren.
Eiwit-eiwit interacties
De laatste manier waarop transductie van hydrofiele signalen kan plaatsvinden is via eiwit-eiwit
interacties. In het plaatje hiernaast zie je twee inactieve RTK’s. Deze RTK-moleculen diffunderen vrij
door het celmembraan. Wanneer er een signaalmolecuul zich aan twee RTK’s hecht, zal de transfosforylatie plaatsvinden. Fosfaateenheden kunnen
zich aan kinase-eenheden binden omdat de
ruimtelijke structuur na het binden van het
signaalmolecuul veranderd is.
Zoals je kunt zien heeft elk RTK-molecuul een
kinase eenheid. Deze worden geactiveerd doordat
een signaalmolecuul zich aan twee RTK eenheden
hecht. Hierna begint de echte trans-fosforylatie.
Na de trans-fosforylatie kunnen verschillende
eenheden zich aan deze fosfaatgroepen binden,
zoals SH2 en SH3 moleculen. Verder zie dia’s.
Groeifactor via RAS (speelt o.a. een rol bij apoptose):
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
7
Celbiologie 12 september 2011
dr. P van Haastert
Het cytoskelet
Er zijn in de celbiologie 3 typen eiwit filamenten te herkennen. Ten eerste zijn er de actine
filamenten. Deze zorgen vooral voor de bewegelijkheid van de cel. Ten tweede zijn er de microtubuli,
deze zorgen voor een soort transportwegen in de cel. Tenslotte zijn er nog de intermediaire
filamenten. Deze zorgen er voornamelijk voor dat de cel bij elkaar wordt gehouden.
Actine filamenten
Actine is een eiwit dat onderdeel is van het cytoskelet in de
eukaryote cel. Actinemoleculen maken onderdeel uit van
actinefilamenten (ook wel microfilamenten genoemd), een
polymeer van actinemonomeren. Ze geven de cel vorm en
maken het mogelijk dat ze de spanning van de cel kunnen
‘dragen’. In spiercellen zorgen ze samen met myosine voor
samentrekking, waardoor spieren kracht uit kunnen oefenen. Bij
de deling van een dierlijke cel spelen actine en myosine een rol
bij de insnoering van de cel tot twee nieuwe. In het plaatje
hiernaast zijn de actine filamenten rood, en de microtubuli
groen gekleurd.
Het is erg belangrijk dat filamenten in een cel snel afgebroken
en opgebouwd kunnen worden (zie plaatje hiernaast). Zodra er
een signaal de cel binnenkomt, zullen de filamenten in het linker deel van de cel afbreken en zullen
er rechts de benodigde nieuwe gevormd worden.
De stabiliteit van een filament hangt af van het aantal
bindingen dat een subunit van zo’n filament is
aangegaan. In het plaatje hiernaast zie je een voorbeeld.
Het is vrijwel onmogelijk om zo’n filament te breken. De
snelheid van de vorming van zo’n filament hangt af van
de grootte van het filament bij het starten van de
polymerisatie (zie dia’s). Zodra je begint met een groter
‘molecuul’, zal je sneller een groter filament hebben dan
wanneer je met één subunit begint te polymeriseren.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
8
Intermediaire filamenten
De intermediare filamenten bestaan uit polymeren
van eiwitten die specifiek zijn voor celtypen, met
een diameter van 8-12 nanometer. Zo bestaan deze
filamenten bij neuronen uit andere eiwitten dan
bijvoorbeeld in rode bloedcellen. Ze verrichten
verschillende functies, afhankelijk van het celtype.
Evenals de actine filamenten dragen de
intermediaire filamenten de spanning van de cel
(zie plaatje hiernaast).
Microtubuli
De microtubuli zijn buisachtige structuren met een dikte van ongeveer 25 nanometer. Microtubuli
zijn opgebouwd uit ketens van tubulinedimeren die samen een buis vormen. Microtubili zijn in
principe opgebouwd uit ringen. Deze ringen bestaan uit ieder 13 monomeren. Deze filamenten zijn
meestal aan 'één kant verbonden aan het centrosoom, van waaruit verschillende tubuli in alle
richtingen uitstralen. Microtubuli zijn belangrijk bij de celdeling, waar ze de chromosomen uit elkaar
trekken. Ook zijn ze erg belangrijk voor het transport van organellen door de cel. Hieronder een lijstje
met verschillende eigenschappen van microtubuli
1. opbouw uit asymetrisch alfa
beta-tubuline dimeer.
2. microtubulus filament is ook
asymetrisch.
3. groei en afbraak aan + kant; kant stabiel.
4. geen afbraak midden in
filament.
5. de beta-tubuline bindt GTP en
is GTPase.
6. filament is stabiel met GTP aan
+ uiteinde,
waardoor meer polymerisatie.
7. filament is instabiel met GDP
aan + uiteinde,
waardoor filament afbrokkelt.
8. gevolg is groei en catastrofe.
Het langer worden van een microtubule
GTP zorgt ervoor dat de dimeereenheden (alfa,beta-tubuline
dimeer) blijven ‘plakken’ aan de vormende streng. GTP is dus
niet nodig voor de polymerisatiereactie zelf. Na een tijdje zal
de GTP in de streng veranderen in GDP (zie onderste streng v/d
linker microtubule hiernaast). Dit geeft niet omdat de streng
inmiddels een stuk verder is gepolymeriseerd. Mocht dit
gebeuren op de plek waar de tubule op dat moment aan het
polymeriseren is, dan brokkelen de filamenten af (rechter
situatie).
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
9
Aan de - kant (aan de + kant vinden alle groei en afbraakreacties
plaats) wordt de microtubule vastgehouden door zogenaamd
gamma tubuline. Als je goed kijkt in het plaatje op de eerste
pagina, zie je dat veel microtubuli vanaf de kern, richting het
celmembraan lopen.
Zogenaamde motoreiwitten zijn in staat organellen te
transporteren van + naar - en van - naar +. Het is dus belangrijk dat
je onthoudt dat microtubuli van - naar + groeien. Het eiwit Kinesine
gaat meestal van de - naar de + kant, en Dyneine van + naar -.
Zodra ze het einde van de streng bereiken, zullen ze losraken van
de ‘rails (dus de microtubule)’ en losraken in het cytoplasma waar ze weer opnieuw kunnen worden
gebruikt.
Microtubuli zijn erg belangrijk in de cel. Zo wordt bijvoorbeeld de structuur van het Golgi apparaat bij
elkaar gehouden door microtubuli. Ook trekken ze tijdens de celdeling (anafase) de chromosomen uit
elkaar. Om overmatige celdeling (=kanker) tegen te gaan, worden er stoffen (chemo op taxolbasis) in
de bloedbaan van de patient gebracht, die ervoor zorgen dat de microtubuli niet meer in staat zijn
om de chromosomen uit elkaar te trekken. Hierdoor zullen beide dochtercellen sterven, en de kanker
dus geremd worden.
Flagel en cilia (trilhaartjes in longen)
Flagella zijn lange staartvormige structuren, cilia zijn korter. Ze bestaan voornamelijk uit tubuline en
kunnen gebruikt worden bij de voortbeweging, bij het binnenhalen van voedsel of bij aftasten van de
omgeving. Bij eukaryoten bestaat een flagel uit een uitstulping van het celmembraan. Het centrum
bestaat uit een axoneem, een cilinder van 9 paren microtubuli met nog 2 microtubuli in de kern. De
basis van die koker wordt gevormd door het
basale lichaampje. Dit is opgebouwd uit een
cilinder van negen tripletten microtubuli, analoog
aan een kern. Het motoreiwit dyneïne beweegt
de microtubuliparen van het axoneem ten
opzichte van elkaar. Omdat ze onderling
verbonden zijn met linking eiwitten, nexin in dit
geval (en dus niet ten opzichte van elkaar kunnen
schuiven) buigt het flagel. Hierdoor ontstaat een
slagbeweging die de cel in een richting loodrecht
op de as van de flagel doet voortbewegen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
10
Intermediaire filamenten
Intermediaire filamenten zijn enkel aanwezig in cellen die een meercellige organisatie vertonen. Een
essentiële rol van intermediaire filamenten is het verdelen van trekkrachten over cellen in een
weefsel. In tegenstelling tot microtubuli en microfilamenten (actine eenheden) worden intermediaire
filamenten gevormd uit een groot aantal verschillende proteïnen. Hoewel intermediaire filamenten
veel stabieler zijn dan
microfilamenten en microtubuli,
wisselen ze gemakkelijk subeenheden
uit van een opgeloste poel.
Omdat een alfa monomeer (keratine),
hydrofobe en hydrofiele kanten
heeft, zullen twee
monomeereenheden om elkaar
draaien en een helix vormen. Ook
deze helices zullen symmetrisch naast
elkaar gaan liggen, zodat er een
lange, symmetrische keten zal
ontstaan.
Werkingsmechanisme actine
Ze geven de cel vorm en maken het
mogelijk dat de cel zich kan bewegen.
In spiercellen zorgen ze samen met
myosine voor samentrekking,
waardoor spieren kracht uit kunnen oefenen. Bij de deling van een
dierlijke cel spelen actine en myosine een rol bij de insnoering van de
cel tot twee nieuwe.
Enkele eigenschappen van actine:
- Actine asymmetrisch
- Actine filament ook asymetrisch; +kant en –kant
- +kant = barbed end = kant van snelste polymerisatie
- Vele actine bindende eiwitten die:
- Polymerisatie actine beinvloeden
- Nucleatie kern voor polymerisatie zijn
- Filamenten cross-linken
- Actine filament aan membraan hechten
- Over filament bewegen (myosine)
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
11
Start actine filament
Net als microtubuli hebben actinemoleculen
ook een + en een - kant. In het plaatje
hiernaast kun je zien hoe de polymerisatie
verder verloopt.
Spierwerking
De samentrekking van spieren is mogelijk door
twee eiwitten: actine en myosine. Deze twee
eiwitten vormen vezels die in elkaar kunnen
schuiven. De binding tussen actine en myosine
is bijzonder sterk en wordt op verschillende
manieren in de bio-technologie gebruikt. Deze schuifactie kost energie en wordt door ATP en GTP
gevoed. Voor verdere informatie moet je maar even in de dia’s kijken, die plaatjes spreken wel voor
zich. Hieronder nog een weergave van hoe actine langs myosine schuift.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
12
Celbiologie 12 september 2011
dr. P van Haastert
Regulatie van celdeling
Alle organismen (één EN meercelligen) zijn het product van cel groei en celdeling. De cyclus waarin
de cel zijn inhoud dupliceert en deelt heet de celcyclus. Afwijkingen aan deze celcyclus kan leiden tot
vergroting van organen, of zelfs tot kanker.
Mitose
Profase
Na de interfase waarin het erfelijk materiaal verdubbeld is volgt de eerste fase van de mitose: de
profase of voorbereidingsfase. De profase wordt gekenmerkt door:
vorming van de spoelfiguur
verdwijnen van de kernmembraan
spiralisatie van en aanhechting van de chromosomen.
In eukaryote cellen ligt net buiten de kernmembraan het centrosoom, een gebied waarin bij dierlijke
cellen de twee centriolen liggen. De centriolen verdubbelen zich in het begin van de profase en
bewegen zich dan elk langzaam naar één kant van de kern. Geleidelijk ontwikkelt zich uit elk van de
centrosomen een structuur van draden. Deze
draden zijn gemaakt van microtubuli (Latijn:
tubuli, buisjes). Een aantal evenwijdig lopende
microtubuli vormt samen bundels,
spoeldraden genaamd, die groot genoeg zijn
om met de lichtmicroscoop te zien. De
microtubuli zijn gemaakt van het eiwit
tubuline.
Metafase
Een cel met een spoelfiguur heeft twee
uiteinden, de polen, en in het midden het
equatoriaalvlak. Wanneer de centromeren van
elk van de chromosomen in één vlak in het
centrum (equatorvlak) van de cel liggen,
begint de metafase. De metafase is afgelopen
zodra de chromatiden loslaten, doordat het
centromeer zich in tweeën deelt. De twee
chromatiden vormen nu elk een zelfstandig
chromosoom.De metafase is de fase waarin de
chromosomen het duidelijkst bestudeerd
kunnen worden, deze chromosomen heten
metafase-chromosomen. De metafase duurt
ongeveer 20 minuten. Het is het langste
onderdeel van de mitose
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
13
Anafase
De beslissende fase van de mitose is de anafase. Het is alsof na een signaal de verbindingen tussen
de chromatiden, op de plaats van de centromeren, loslaten en de enkele chromatiden (nu
chromosomen) zich elk in tegenstelde richting naar de polen bewegen doordat de spoeldraden deze
naar de polen trekken. Dit gebeurt omdat de trekdraden verkorten. De anafase stopt wanneer elke
set chromosomen aangekomen is aan zijn pool. Hier zullen de nieuwe kernen gevormd worden. De
anafase is de kortste fase van de mitose. Naar welke pool de chromatiden zich begeven, is
willekeurig. De fase duurt hooguit een paar minuten.
Telofase
Tijdens de telofase despiraliseren de chromosomen , ontrollen zich, en worden weer lange dunne
draden, die niet van elkaar te onderscheiden zijn met lichtmicroscoop. De chromosomen worden
opnieuw 'verpakt' in een kernenvelop en de vorming van twee complete kernen is dan voltooid.
Nadat de telofase ten einde is gekomen, is het resultaat twee nieuwe kernen. Hiermee is de mitose
beëindigd. Nu volgt de cytokinese of celdeling.
Fasen van de celcyclus
Eukaryote cellen hebben eenzelfde celcyclus, deze is
ruwweg in te delen in twee fases, namelijk:
Interfase, voorbereiden op het delen van de
cel.
Mitose of meiose, de uiteindelijke deling van de
kern.
De Interfase wordt ingedeeld in drie fases:
De G1-fase, belangrijke toename van
cytoplasma in de cel. En het aanmaken van
eiwitten, ter voorbereiding op de celdeling.
De S- of synthesefase, de cel gaat alle chromosomen of anders gezegd alle informatie die de
kern bevat voor het functioneren van de cel kopiëren door replicatie. Het materiaal kan dan
bij de mitose worden gedeeld. Elk van de twee nieuwe chromosomen heet nu chromatide
De G2-fase, de periode tussen het verdubbelen van het DNA en de deling van de cel. In deze
fase worden stoffen gemaakt die belangrijk zijn voor de celdeling.
Tussen G1 en S fase en na de G2-fase zijn er ook 'checkpoints' (controlepunten). CH2
(checkpoint 2, na G2)controleert of de DNA-replicatie goed is gebeurd. Kleine fouten worden
hersteld, maar als er zich te grote fouten voordoen gaat de cel in apoptose
(geprogrammeerde celdood)
Karakteristieken van het celcyclussysteem
Zoals je op het plaatje boven aan deze pagina kan zien, worden de verschillende fasen in
chronologische volgorde doorlopen, dit gebeurd altijd! Een cel kan nooit een fase overslaan. De ene
fase zal langer duren dan de andere, maar elke fase wordt doorlopen tijdens een cyclus. Na elke fase
zit een soort ‘stop’, deze stop wordt opgeheven wanneer de laatste fase helemaal afgelopen is
(bijvoorbeeld nadat het laatste chromosoom aangehecht is aan een microtubule). Pas dan zal de
volgende fase van de mitose beginnen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
14
Controle van de celcyclus
Omdat de celcyclus een heel complex systeem is, dient het ook
gecontroleerd te worden. De celcyclus kent 3 belangrijke
controlepunten.
Elk controlepunt heeft twee belangrijke componenten: een
cycline-eiwit en een cycline-afhankelijke base. Deze eiwitten zijn
geprogrammeerd om alleen actief te zijn tijdens een bepaalde
fase van de celcyclus. Na hun taak worden cycline-Cdk’s inactief,
omdat de cycline eenheden worden afgebroken.
De cycline-Cdk concentraties stijgen telkens voordat
de G1-, de S- en de G2-fase beginnen en vallen
drastisch terug aan het einde ervan. Men neemt aan
dat een controlepunt bereikt wordt op het moment
dat de maximum hoeveelheid van deze cycline-CdK's
aanwezig is. Daarna vindt weer een snelle afbraak van de cycline-CdK's tot cyclines en Cdk's plaats.
De CdK's fosforyliseren een groep andere eiwitten en sturen zo de celcyclus. Hieronder is
weergegeven hoe inactieve Cdk’s (in dit geval M-Cdk, deze stof zorgt er voor de overgang naar de Mfase (mitose)).
Verschillende typen Cdk:
Cycline D - cdk4 complex : werkt gedurende het midden van de G1 fase, Dit is het restrictiepunt
(R), een cruciaal beslissingspunt dat eens gepasseerd de volledige celcyclus laat doorlopen.
Cycline E - cdk2 complex : werkt ook in het midden van de G1 fase, G1-S overgang.
Cycline A - cdk2 complex : werkt gedurende de S fase en stimuleert de DNA replicatie, S-G2
overgang.
Cycline B - cdk1 complex : werkt ter hoogte van de G2-M overgang en initieert de Mitose
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
15
Apoptose
Dit proces is essentieel tijdens de ontwikkeling en voor het in stand houden van weefsels in de adult
(celdeling ± 1 kg per dag). Ook is apoptose essentieel voor het verwijderen van niet-functionele
cellen van het immuunsysteem. Het doel van apoptose is dus ook het doodmaken van een cel,
zonder het organisme te beschadigen (dit i.t.t. necrose, het
openbreken van cellen, waardoor er een immuunrespons
optreedt).
Caspase
Een caspase is een lid van een groep cysteïneproteasen,
enzymen met een cruciale cysteïnegroep die andere eiwitten
kunnen klieven na een aspartaatgroep. Caspasen vervullen in
de cel een essentiële rol bij de apoptose, een van de
belangrijkste mechanismen voor geprogrammeerde celdood
tijdens de ontwikkeling en gedurende de meeste stadia van het
volwassen leven. Caspasen zijn wel de 'beul-eiwitten' genoemd
vanwege deze rol in de cel.
In een cel bevinden zich standaard inactieve procaspases. Deze
procaspases zijn een beetje actief. Uit zichzelf zullen ze nooit
opeens overgaan in een actieve caspase. Dit gebeurt pas
wanneer er veel beetje actieve- procaspases bij elkaar komen.
Ook zal een procaspase overgaan in een caspase wanneer deze
in aanraking komt met een actief-caspasemolecuul. Zo ontstaat
er een lawine van actieve caspasemoleculen die de cel zullen
afbreken.
Apoptose in een witte bloedcel
Hieronder een voorbeeld van hoe een killerlymfocyt een inactieve witte bloedcel tot apoptose
aanzet. Hier koppelt de k-lymfocyt zich aan de target cel met een bepaald koppeleiwit die precies
past op de Fas death receptor. Hierdoor zullen moleculen zich aan de binnenkant van het
celmembraan hechten, waardoor er veel procaspase-moleculen dicht bij elkaar komen. Hierdoor
zullen ze omgezet worden in een actief-caspasemolecuul, die de cel zal afbreken.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
16
Apoptose in mitochondriën
Ook in organellen zoals mitochondrien is het belangrijk dat alles goed verloopt. Zodra een
mitochondrion niet meer goed werkt, of lek is, zullen er cytochromen naar buiten lopen die weer een
cascade van reacties zal veroorzaken wat uiteindelijk zal leiden tot apoptose van de cel (zie
voorbeeld hieronder). Ook hier zullen inactieve procaspases omgezet worden tot caspases die de cel
zullen gaan ‘slopen’.
Een mitochondrion kan zich ook tegen
actieve caspases weren. Zo kunnen Bcl2moleculen op samengeklonterde poriën in
het membraan van een mitochondrion gaan
zitten, zodat er cytochromen naar buiten
kunnen (wat dus zou leiden tot apoptose).
Wanneer er een apoptotische stimulus van buiten komt, zal het Bcl2 molecuul van de poriën afgaan,
zodat er toch cytochromen naar buiten zullen lopen. Dit kan je zien als een soort commando van de
cel. Ook kunnen er IAP’s gemaakt worden, die actieve caspasemoleculen zullen inhiberen (en dus
onschadelijk maken). Zoals je in het plaatje hiernaast kunt zien, krijgt een cel constant
overlevingssignalen binnen die er voor zorgen dat de cel in leven blijft. Deze signalen zetten de
volgende reacties in werking. Deze reacties vormen dus die Bcl2 en IAP-moleculen die er voor zorgen
dat caspasemoleculen niet de kans
krijgen om de cel kapot te maken.
Er zit een bepaalde hiërarchie in
deze reacties, de één zal dus
sterker zijn dan de andere. Eén ding
is zeker, aan één of twee van de
drie overlevingssignalen heeft de
cel niet genoeg om in leven te
blijven.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
17
Celbiologie 19 september 2011
dr. I. van der Klei
Endoplasmatisch reticulum
Het endoplasmatisch reticulum is een netwerk
(reticulum) van membranen dat gelegen is in het
cytoplasma van een cel. Het bestaat uit twee
dicht tegen elkaar liggende membranen
waartussen holten en kanalen worden gevormd.
Het is afgescheiden van de rest van de cel door
een membraan met dezelfde structuur als het
celmembraan.
Ruw ER
Het ruw endoplasmatisch reticulum (RER of
ribosomaal ER) herbergt de ribosomen en heeft
daardoor een belangrijke rol in de eiwitvorming
(eiwitsynthese) in de cel. Ook heeft het een rol in
het transport van stoffen in de cel, met name is
het belangrijk voor het verzamelen van eiwitten
die naar het golgi-apparaat vervoerd moeten worden. Het ruw endoplasmatisch reticulum dankt zijn
naam aan het ruwe uiterlijk onder de elektronenmicroscoop, dat veroorzaakt wordt door de vele
ribosomen die op het buitenoppervlak zijn aangehecht. Alle aangehechte ribosomen zijn verbonden
met de binnenzijde van het ER door een porie-eiwit. Wanneer een van deze ribosomen een mRNAstreng ontvangt, wordt de door translatie gevormde peptideketen via het porie-eiwit het ER ingeleid.
Pas daarna wordt het eiwit tot zijn uiteindelijke vorm gevouwen.
Glad ER
Het glad ER is een variant van ER die voornamelijk dient om stoffen vanuit het ruw ER te vervoeren
naar het golgi-apparaat, het bevindt zich dan ook vaak tussen zones met ruw ER en het golgiapparaat. Andere functies van het glad endoplasmatisch reticulum zijn het opslaan van calciumionen
en de synthese van lipiden.
Golgi apparaat
In het golgi-apparaat worden de producten afkomstig van het endoplasmatisch reticulum (ER)
omgebouwd en opgeslagen, om dan later naar andere bestemmingen verscheept te worden. Het is
niet verwonderlijk dat vooral de cellen van secretie-organen bijzonder veel golgi-apparaten bezitten.
Het golgi-apparaat bestaat uit een stapel platte cisternen met enige ruimte ertussen. De cisternen
zijn met hun cis-kant richting celkern gekeerd en met hun trans-kant richting het buitenste
celmembraan en dus de wereld buiten de cel, omdat daar blaasjes met omgebouwde producten
ontstaan en zo de cel via het cytoplasma zullen willen verlaten.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
18
Vacuole en lysosoom
Het lysosoom is een blaasje dat zich in het cytoplasma bevindt
en ook wel "de afvalberg van de cel" genoemd wordt.
Lysosomale enzymen in de lysosomen breken afvalstoffen van
de cel af, zodat de afbraakproducten hergebruikt of veilig
uitgescheiden kunnen worden. In planten wordt een lysosoom
ook wel de vacuole genoemd. De onverteerde voedingsstoffen
worden aangeleverd door endosomen en fagosomen. In het
secundaire lysosoom heerst een zuur milieu met een pH van
rond de 5. Dit is nodig omdat lysosomale enzymen alleen
werken bij zo’n lage pH. Een lysosoom houdt zijn ph laag
doormiddel van een H-pomp. Hierbij wordt een ATP omgezet tot
ADP+P. De energie die hierbij vrijkomt wordt gebruikt om een
H+-molecuul naar binnen te pompen, waardoor de pH in het
lysosoom daalt.
Peroxisomen
Een peroxisoom (ook: peroxysoom) is een celorganel in een eukaryote cel. Een enkel membraan sluit
hem af van het cytosol (interne vloeistof van een cel). De functie van peroxisomen is het lichaam te
ontdoen van toxische substanties zoals waterstofperoxide en andere metabolieten. Zij komen bijna
in elke cel van het lichaam voor; grote hoeveelheden peroxisomen treft men aan in de cellen van de
lever waar toxische bijproducten worden gestapeld.
Peroxisomen zijn in staat waterstofperoxide (H2O2) te
vormen door waterstof bij zuurstof te plaatsen. Het
peroxisoom is DE plaats voor de zeer langketen vetzuur
afbraak in de meeste cellen. VLCFA's of very long chain
fatty acids zijn zeer langket en vetzuren met een koolstof
ketenlengte van meer dan 22 koolstofatomen; de
peroxisomale β-oxidatie verkort de ketenlengte totdat de
VLCFA's verder afgebroken kunnen worden in de
mitochondriën. Onder de
microscoop zijn peroxisomen te
herkennen aan de kristallen in de
matrix van het peroxisoom. Dit zijn
allemaal dezelfde eiwitten.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
19
Microscopie
Lichtmicroscopen of optische microscopen
De grens voor optische microscopen ligt bij ongeveer 2000×; sterkere vergrotingen zijn wel mogelijk,
maar laten niet meer details zien. Zichtbaar licht heeft een golflengte tussen 380 en 780 nm. Details
die kleiner zijn, zijn niet zichtbaar. Deze microscopen worden vooral in de geneeskunde en de
biologie gebruikt, voor het bekijken van micro-organismen, cellen en weefsels. Biologische
microscopen hebben vaak twee oculairen, zodat men tegelijkertijd met beide ogen („binoculair”) kan
kijken. Men ziet echter geen diepte, omdat beide ogen door hetzelfde objectief kijken.
Fasecontrastmicroscopen
De fasecontrastmicroscoop lijkt veel op een
lichtmicroscoop maar is voorzien van een speciaal
‘faseplaatje’ tussen de condensor en het preparaat. Dit
zorgt ervoor dat twee lichtbundels met een onderling
faseverschil door het transparante preparaat vallen.
Door interferentie van deze twee beelden worden
vrijwel transparente voorwerpen (cellen e.d.) zichtbaar
zonder dat er een kleuring nodig is. Ook ontstaat enig
diepte-effect, doordat deze voorwerpen donkerder
lijken naarmate de dikte groter is.
Toch blijven er beperkingen. Ondanks alle verbeteringen op het gebied van microscopie is het nog
steeds lastig om een goed contrast te creëren in ‘levende’ weefsels. Ook moeten er plakjes van het
originele preparaat worden gesneden, om het te kunnen bekijken onder de microscoop. Hierdoor zal
het preparaat altijd dood of stervende zijn. Ook moet er aan een preparaat voor de lichtmicroscoop
ook een kleuring worden toegevoegd (dit moet bij vrijwel alle
dierlijke cellen).
Fluorescentiemicroscopie
Fluorescentie is een bijzonder geval van luminescentie. Het is een
natuurkundig verschijnsel waarbij een atoom een hoogenergetisch foton absorbeert, in een aangeslagen toestand
belandt en vervolgens terugvalt naar de grondtoestand onder
uitzending van een foton van lagere energie (langere golflengte).
Fluorescentiemicroscopie is een techniek die in biologisch en
medisch onderzoek wordt gebruikt waarbij fluorescerende
kleurstoffen worden gebruikt die oplichten als ze worden bestraald met licht van een kortere
golflengte. De meeste vormen van fluorescentiemicroscopie maken gebruik van sera van kunstmatig
geproduceerde antistoffen. De methode wordt dan ook wel immunofluorescentie-, ofwel IFmicroscopie genoemd. Deze zijn afkomstig van bijvoorbeeld muizen of ratten, en zijn specifiek
gericht tegen een bepaald, bekend eiwit. Als het serum wordt toegevoegd aan een weefsel, zullen de
antistoffen binden aan het doeleiwit. Niet-gebonden antistoffen worden weggewassen. De
antistoffen zijn voorzien van een fluorescente kleurstof, en daarmee worden ze microscopisch
zichtbaar.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
20
Werking Fluorescentiemicroscopie
Met een fluorescentiemicroscoop kunnen mooie plaatjes worden
gemaakt. Het is echter maar mogelijk om met 1 kleur tegelijk te
werken. Plaatjes zoals hiernaast worden gemaakt door eerst bijv. GFP
te gebruiken en daarna een rode kleuring. Van al deze stappen wordt
een foto gemaakt, je hebt dus een groene, rode en blauwe foto. Deze
foto’s worden later over elkaar geplakt met photoshop om zo’n
plaatje als hiernaast te krijgen.
Immunofluorescentie
Bij immunofluorescentie wordt gebruik gemaakt van een antilichaam
dat specifiek aan het te onderzoeken eiwit (X) bindt. Wanneer dit antilichaam (A) wordt toegevoegd
aan het te onderzoeken monster, hecht deze zich vast aan het specifieke eiwit (X) indien dit aanwezig
is. Na wegspoelen van niet gebonden antilichamen blijven alleen de gekoppelde antilichamen (A)
over. Vervolgens worden antilichamen (B) met een fluorescerende groep toegevoegd, die specifiek
binden aan A. Na wegspoelen van de niet gebonden antilichamen (B) kan het monster worden
bekeken met een fluorescentiemicroscoop. De fluorescerende groepen van de antistoffen (B) zullen
nu oplichten. Op deze manier kan aangetoond worden of de eiwitten aanwezig
zijn en zo ja, waar de eiwitten zich bevinden. In de afbeelding hieronder is
gewerkt met een antigen i.p.v. een eiwit.
Confoca
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
21
le laser scanning microscoop
Bij confocale laserfluorescentiemicroscopie wordt gebruikgemaakt van de laser scan microscoop
(LSM). Het principe van de confocale laserscanmicroscopie is begin jaren '50 ontwikkeld en in 1957
gepatenteerd door de Amerikaanse wetenschapper Marvin Minsky. Pas nadat halverwege de jaren
'80 laserlicht gebruikt werd, heeft fluorescentiemicroscopie een grote vlucht genomen.
Het meest gebruikte type LSM van vandaag de dag is de confocale laser scan microscope (CLSM). In
dit apparaat zijn een aantal kenmerkende onderdelen te vinden:
laser; straalt monochromatisch licht uit.
halfdoorlatende spiegel; reflecteert een deel van het licht, maar laat de andere helft door
pinhole (speldeprikgrote opening); blokkeert ongewenst licht
detector; zet licht om in elektrisch signaal
Via de halfdoorlatende spiegel en een serie lenzen bereikt het licht van de laser het preparaat. Het
als gevolg van de fluorescentie geëmitteerde licht komt via de lenzen en de spiegel terecht bij het
"pinhole" met direct daarachter de detector (een fotomultiplicator). Het pinhole zorgt ervoor dat
alleen het licht van een zeer dunne plak van het preparaat (het focale vlak) de detector bereikt.
Fotonen van buiten het focale vlak worden door het pinhole geblokkeerd. De detector is verbonden
met een computer zodat digitale beelden direct verwerkt en geanalyseerd kunnen worden. De op
deze manier verkregen beelden zijn veel scherper dan beelden van conventionele microscopen.
Door de spiegel te bewegen kan de positie (X/Y) van de laserstraal op het preparaat worden bepaald.
Met de pinhole kan de hoogte (Z) worden bepaald. Door een serie opnamen te maken waarbij de
driedimensionale coördinaten in het preparaat afzonderlijk een voor een worden afgetast, en deze
met behulp van computertechniek te combineren, kunnen driedimensionale beelden van levende
cellen worden gemaakt met een zeer hoge resolutie, meer dan met conventionele optische middelen
mogelijk is.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
22
GFP
Groen fluorescent proteïne of GFP (Green Fluorescent Protein in het Engels) is een fluorescent eiwit
dat voor het eerst in 1962 bij de kwal Aequorea victoria werd ontdekt. Er zijn veel verschillende
varianten van GFP ontwikkeld, die bijvoorbeeld in plaats van groen een andere kleur licht uitzenden,
of die chemisch stabieler zijn.
Fluorescerende eiwitten hebben een zeer belangrijke rol gekregen in de moleculaire biologie, waar
ze vooral gebruikt worden om andere eiwitten te labelen. Met behulp van fluorescentiemicroscopie
kan men dan bijvoorbeeld traceren waar in de cel een eiwit zich bevindt. Fluorescente labeling is
veiliger en gevoeliger dan radioactieve labeling, wat voorheen veel gebruikt werd.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
23
Celbiologie 20 september 2011
dr. I. van der Klei
Elektronenmicroscoop
Elektronenmicroscopie is een techniek die gebruik
maakt van een bundel elektronen om het oppervlak
of de inhoud van objecten af te beelden. Doordat
versnelde elektronen een veel kleinere golflengte
hebben dan fotonen kan de resolutie van een
elektronenmicroscoop veel hoger zijn (beter dan 0,1
nm) dan die van een lichtmicroscoop (ongeveer 200 nm).
Daarnaast hebben elektronen een andere wisselwerking met de materie zodat er een ander contrast
verkregen kan worden. Bij lichtmicroscopie wordt de resolutie beperkt door de golflengte van het
licht, bij elektronenmicroscopen wordt de resolutie beperkt door de afwijkingen van de optiek, want
elektronenlenzen zijn in vergelijking met optische lenzen van veel slechtere kwaliteit.
Omdat je met elektronenmicroscopen altijd gebruik moet maken van splinterdunne preparaten, is
het noodzakelijk dat deze preparaten met een machine gemaakt worden. Er wordt in een
elektronenmicroscoop altijd gewerkt in een vacuüm, zodat de elektronen vrij kunnen bewegen. Om
te zorgen dat dit vacuüm het preparaat niet beïnvloedt, voegt men chemische fixatieven toe,
waardoor het preparaat hard wordt.
Scanning EM
Een rasterelektronenmicroscoop (Engels: Scanning EM) werkt door
middel van een zeer fijne, scherp gefocusseerde elektronenbundel die
versneld wordt afgeschoten op het preparaat. De gebruikte
versnelspanning varieert tussen 100 V en 30 kV. De bundel wordt
uitgelijnd door elektromagnetische velden, die in de
elektronenmicroscoop eenzelfde functie vervullen als optische (glazen)
lenzen in een lichtmicroscoop. De bundel wordt door
elektromagnetische velden snel heen en weer bewogen langs lijnen en
na iedere lijn een beetje opgeschoven in de richting loodrecht op de lijn,
op de manier waarop een televisiebeeld wordt opgebouwd. Dit gebeurt
volgens een beeldraster, vandaar ook de naam. Het preparaat wordt zo
gescand. Dit levert mooie, maar onrealistische plaatjes op. Omdat er met
elektronen wordt gewerkt, kun je nooit kleuren zien. Dit is allemaal later
toegevoegd met een computer.
Eiwittransport
We kunnen in de celbiologie 3 soorten transport onderscheiden. Gated,
transmembrane en vesicular transport. Om al deze transportprocessen
goed te laten verlopen, hebben alle eiwitten een signaaldeel die ervoor
zorgt dat het eiwit op de plek komt waar hij moet zijn.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
24
Signaaleiwitten
A signal peptide is a short (3-60 amino acids long) peptide chain that directs the transport of a
protein. Signal peptides may also be called targeting signals, signal sequences, transit peptides, or
localization signals.The amino acid sequences of signal peptides direct proteins (which are
synthesized in the cytosol) to certain organelles such as the nucleus, mitochondrial matrix,
endoplasmic reticulum, chloroplast, apoplast and peroxisome. Some signal peptides are cleaved from
the protein by signal peptidase after the proteins are transported.
Transport door kernporie
De celkern en het cytoplasma zijn gescheiden door de
kernmembraan. De kernmembraan bevat honderden
kernporieën, die de toegangspoorten tot de celkern
vormen. Een kernporie wordt gevormd door het nuclear
pore complex (NPC). Het NPC behoort met een gewicht
van 125 miljoen Dalton (bij zoogdieren) tot de
allergrootste eiwitcomplexen. Het bevat vele tientallen
verschillende eiwitten. Het hart van het NPC wordt
gevormd door 8 grote 'spaken' die in een ringvorm in de
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
25
kernmembraan zitten. Deze spaken zitten ingeklemd tussen twee kleinere 'ringen': een aan de zijde
van het cytoplasma (waaraan een aantal filamenten bevestigd zijn), en een aan de zijde van het
nucleoplasma. Aan de zijde van het kernplasma hangt een 'mandje', dat wordt gevormd door 8
filamenten en een terminale ring. In het midden van het NPC bevindt zich een moeilijk te
identificeren plug. Deze bestaat mogelijk uit eiwitten die door het NPC worden getransporteerd.
Kleine moleculen kunnen vrij door de porie diffunderen. Grote moleculen kunnen dit niet, dit gaat
dan via actief transport met GTP (geen ATP zie onderste afbeelding vorige pagina).
Transport door een kernporie:
- Import en export van eiwitten
- Actief eiwittransport is GTP afhankelijk
- Export RNA moleculen
Verschillende grote eiwitten hebben dus allemaal een
signaaldeel. Op dat signaaldeel past precies een bepaalde
receptor met GDP. Zodra dit complex in de kern matrix komt,
zal Ran-GTP ervoor zorgen dat het eiwit en de receptor
gesplitst worden, door de receptor (met GTP) te fosforyleren
tot GTP. Dit receptor-GTP complex zal de kern weer verlaten.
Buiten de kern zorgt Ran GDP ervoor dat de GTP-groep aan het
receptor-GTP complex omgezet wordt tot een GDP groep. Nu
zijn we weer bij het begin en kan de hele cyclus weer opnieuw
beginnen.
Import naar mitochondriele matrix
Soms moeten eiwitten niet in de celkern geimporteerd worden, maar in de matrix van een
mitochondrion. Ook deze eiwitten hebben een speciale signaalsequentie die ervoor zorgt dat het
eiwit op zijn plek komt. Bij deze eiwitten zit het signaaldeel aan de N-terminus. Dit signaaldeel is een
amfipatische (dwz. Hydrofoob en hydrofiel) alfa helix. Wanneer een eiwit het membraan van een
mitochondrion nadert, hecht het signaaldeel aan de receptor van het TOM-complex. Als dit past,
glijdt het eiwit door het translocatiesignaal richting het Transporter Inner Membrane (TIM23)
complex. Zodra het eiwit het TIM 23-complex is gepasseerd (en dus in de matrix van het
mitochondrion zit), zullen er zogenaamde chaperones (ook wel HSP70 genoemd) aan het eiwit
hechten, die ervoor zorgen dat het
eiwit door het TIM23-complex wordt
getrokken, en dus volledig in de matrix
terecht komt. Zodra dit is gebeurd,
zullen de chaperones het eiwit weer
loslaten en zal de eiwit zich gaan
vouwen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
26
In de afbeelding hieronder is het hele proces schematisch afgebeeld. Het is in principe hetzelfde als
de afbeelding op de vorige pagina. Hier is ook te zien dat het Inner-membraan een lading heeft. Zoals
ik al eerder zei, heeft het signaaldeel van het eiwit dat bestemd is voor de matrix van een
mitochondrion een positieve lading. Ook dit zal helpen om het eiwit van A naar B te krijgen.
Verschillende functies van TIM-complexen
TIM23: Het TIM23 complex zorgt voor de translocatie van matrix-gerichte eiwitten. Het zorgt er dus
voor dat eiwitten van het cytosol, naar de mitochondriale matrix worden verplaatst.
TIM22: Het TIM22 complex zorgt voor de integratie van eiwitten naar IN het inner-membraan.
OXA-complex: Dit complex zorgt voor het transport naar het binnen membraan.
Transport naar thylakoid lumen
Een eiwit dat getransporteerd moet worden naar
het thylakoid lumen heeft twee signaaldelen. Eén
om door het outer en inner membraan te komen
en de andere om door het thylakoid membraan te
komen. Het signaaldeel (delen) zit aan de Nterminus en is amfipatisch (hydrofoob en
hydrofiel).
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
27
Celbiologie 21 september 2011
dr. I. van der Klei
Import naar peroxisomale matrix
Het signaaldeel van het eiwit bestaat uit
3 c-terminale aminozuren, de
zogenaamde PTS1. Het eiwit kan
gevouwen worden geimporteerd, maar
het peroxisoom heeft geen grote porien
zoals de kern envelop. Eiwitten die een
rol spelen bij de import van eiwitten
naar het matrix van peroxisomen heten
peroxins. In de afbeelding hiernaast is
Pex5 de receptor van de PTS1.
Import naar ER lumen
Het signaaldeel vaneen eiwit dat naar de het ER
lumen getransporteerd moet worden zit aan de
N-terminus en bestaat uit hydrofobe
aminozuren die gevolgd worden door geladen
aminozuren (zie tabel vorige samenvatting). De
import naar het ER lumen is co-translationeel.
Dit wil zeggen dat het ribosoom dat het mRNA
moet transleren, zich aan het RER hecht en pas
daar het eiwit begint te transleren (zie
afbeelding hiernaast). Om dit te realiseren is
een zogenaamd SRP (signal recognition particle)
nodig. Dit deeltje zorgt ervoor dat het ribosoom
wacht met het transleren van een mRNA
molecuul totdat deze zich aan het RER heeft
gehecht. Zoals je kunt zien, bindt het SRP eiwit zich in het cytosol aan het ribosoom om zo het
ribosoom-SRP complex te vormen. Wanneer dit complex het RER nadert, zal de SRP eenheid zich
hechten aan een SRP receptoreiwit in het membraan. Zodra dit is gebeurd, zal het eiwit in de protein
translocator zakken, alleen het signaaldeel blijft nog hangen (zie afbeelding ernaast). Het signaaldeel
zal later worden afgeknipt (door een peptidase) en afgebroken worden. Kort samengevat komt het
er op neer dat het SRP het signaaldeel kent, en de receptor op het RER, herkent
het SRP.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
28
Glycolysatie van een eiwit
Het ER heeft ook nog een andere fucntie, namelijk
het glycolyseren van eiwitten. Hierbij wordt er
aan elke ASN (asparagine) een suikergroep
gekoppeld, die later in het golgi apparaat weer
gemodificeerd kan worden. Glycolyseerde
eiwitten kunnen dus wel op meerdere plaatsen in
een cel voorkomen, maar ze zijn altijd in het ER
lumen gemaakt!
Opruimen van eiwitten
Zodra er een fout optreedt in bijvoorbeeld de
vouwing of in het glycolysatieproces, zal het eiwit
worden afgebroken. Dit gaat als volgt: Een
bepaald soort chaperone herkent de foutieve
eiwitten en hecht zich daaraan. Dit chaperoneeiwit complex zal zich naar het membraan
begeven. Hier zal het misgevouwen eiwit het ER verlaten door een ER eiwit translocator met
hulpeiwitten. Vervolgens zit er in het cytosol bepaalde enzymen die de suikergroep van het eiwit
afhalen (efficiënt), de zogenaamde glycanase’s. Vervolgens zal het eiwit worden gemarkeerd en
worden afgebroken.
Regulatie chaperonemoleculen
Zoals je ziet zijn chaperones erg belangrijk in dit proces. Op het moment dat er veel foutieve eiwitten
zijn gevormd, is het dus belangrijk dat er voldoende chaperones aanwezig zijn in het ER lumen. Er
zitten zogenaamde transmembrane protein kinase’s in het membraan van het ER. Zodra deze
eiwitten een foutief eiwit zien, zullen ze een andere kinase opzoeken en zich daar aan hechten.
Vervolgens zal dit groepje van twee kinase’s elkaar gaan fosforyleren. Het complex wat nu is
ontstaan is in staat het mRNA wat codeert voor een bepaald gene regulatory protein te splicen
(d.w.z. de introns eruit knippen). Het ontstaande mRNA wordt getransleerd door ribosomen, wat
resulteert in het zogenaamde gene regulatory protein. Dit proteine zal zich naar de kern begeven, om
daar het DNA wat codeert voor chaperonemoleculen te transleren. Dit chaperone mRNA zal zich
vervolgens weer naar het ER begeven om daar getransleerd te worden tot een chaperonemolecuul.
(zie volgende pagina voor een illustratie van dit proces).
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
29
Vesikel transport
Exocytose is het proces waarbij een cel stoffen afgeeft aan of afscheidt naar het celmembraan of het
extracellulaire milieu. Het blaasje waarmee dit gebeurd wordt ook wel een exosoom genoemd. De af
te scheiden stoffen zijn onder andere proteïnen en lipiden. Endocytose is het opnemen van stoffen in
de cel doordat het door het
celmembraan wordt ingesloten. De
stoffen die de celmembraan door
mogen, worden ingesloten doordat
het celmembraan iets naar binnen
toe trekt, steeds verder, totdat het
uiteindelijk een zelfstandig blaasje of
vesikel is, het endosoom.
Coat eiwitten
Clathrin: Dit eiwit coat vesikels die eiwitten vervoeren die de cel zullen verlaten via exocytose, of de
cel binnenkomen via endocytose
COP I: Dit eiwit coat vesikels die eiwitten vervoeren van de cis-kant van het golgi complex, naar het
RER.
COP II: Dit eiwit coat vesikels die eiwitten vervoeren van het RER naar het golgi apparaat.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
30
Assemblage en disassemblage van de coating eiwitten
Clathrin:
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
31
COPII:
Hier zie je ook dat chaperonemoleculen ongevouwen of misgevouwen eiwitten vasthouden, zodat
deze niet meegezonden worden met het blaasje.
SNARE-eiwitten
De voornaamste rol van SNARE-eiwitten is om de koppeling en versmelding tussen verschillende
membranen en versmelting van cellulaire transportvesikels met het celmembraan of het membraan
van een intracellulair compartiment (zoals een lysosoom) mogelijk te maken.
De SNAREs kunnen worden onderverdeeld in twee categorieën: vesikel-, of v-SNAREs die in de
membraan van transportvesikels worden opgenomen als die zich afsnoeren, en target- of t-SNAREs
die zich bevinden in het membraan dat een vesikel moet gaan opnemen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
32
Fusie vesikel met mebraan:
Hier ook uitgelegd hoe de t-SNARE eiwitten weer worden losgemaakt nadat het vesikel gedocked is.
Retrieval transport
Wanneer er per ongeluk eiwitten naar het golgi apparaat zijn verzonden die er niet thuishoren,
worden ze retour gezonden. Dit heeft natuurlijk als eerste functie om het verloren eiwit terug naar
het ER te brengen, maar het heeft nog een functie. Omdat het ER steeds vesikels maakt, krimpt het
langzaam. Om dit proces tegen te gaan, stuurt het ER blaasjes retour.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
33
Het golgi apparaat
Transportblaasjes met eiwitten gemaakt
door ribosomen op het ruw
endoplasmatisch reticulum worden
vervoerd naar het golgi-apparaat, waar ze
hun eiwitten afleveren aan het lumen (de
ruimte binnen het golgi-apparaat). De
eiwitten worden door de membranen
getransporteerd en op hun weg veranderd
in een eindproduct. Hoe dit transport precies werkt is pas kort geleden ontdekt. Het golgi-apparaat
vormt zelf nieuwe cisternen en verwijdert de oude waardoor de eiwitten in de oude cisternen
vrijkomen. Als de eiwitten de trans-kant bereiken, vormen ze blaasjes die, als ze een bepaalde
grootte bereiken, als secreetkorrels richting het celmembraan getransporteerd worden via
microtubili en -filamenten. Het golgi-apparaat maakt ook de
lysosomen aan. Dit zijn blaasjes die partikels, ingenomen door
fagocytose of pinocytose, of oudere celdelen verteren, zodat
de bouwstoffen voor andere zaken kunnen gebruikt worden.
De Cis kant van het golgi apparaat ligt aan de kant van de kern
en de trans kant richting het celmembraan. Tussen de Cis en
de trans kant, liggen de cis cisterna, medial cisterna en de
trans cistena. Dit moet je kennen voor het tentamen!
Zoals ik al zei worden in het golgi apparaat ook glycolyseerde
eiwitten gemodificeerd. Om dit te doen, zitten er andere
enzymen in de verschillende cisternae’s (zie afbeelding
hiernaast).
Opname LDL d.m.v. endocytose
(In de onderste situatie (B) is er een mutatie opgetreden in het LDL
molecuul.)
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
34
Autofagosoom
In cell biology, autophagy, or autophagocytosis, is a catabolic process involving the degradation of a
cell's own components through the lysosomal machinery. It is a tightly-regulated process that plays a
normal part in cell growth, development, and homeostasis, helping to maintain a balance between
the synthesis, degradation, and subsequent recycling of cellular products. It is a major mechanism by
which a starving cell reallocates nutrients from unnecessary processes to more-essential processes.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
35
Celbiologie 22 september 2011
dr. I. van der Klei
Secretie/exocytose
Exocytose is het proces waarbij een cel
stoffen afgeeft aan of afscheidt naar het
celmembraan of het extracellulaire milieu.
De af te scheiden stoffen zijn onder andere
proteïnen en lipiden.
Dit proces loopt via organellen. Te weten:
Endoplasmatisch reticulum
Golgi-apparaat
De af te geven stoffen worden aan het golgiapparaat verpakt in blaasjes (vesikels). De
vesikels worden naar het celmembraan
getransporteerd en fuseren daarmee. Veel verschillende stoffen zijn betrokken bij de exocytose. De
best beschreven stof is de oplosbare NSF receptor (SNARE-eiwitten), die als katalysator werkt bij de
fusiereactie. In de afbeelding hierboven is een constitutieve uitscheiding en een gereguleerde
uitscheiding te onderscheiden. De constitutieve uitscheiding loopt continu, terwijl de gereguleerde
uitscheiding pas gaat lopen zodra er contact wordt gemaakt met signaaleiwit, bijvoorbeeld een
hormoon.
Secretieblaasjes
Om te zorgen dat de hele exocytose goed verloopt, is er een speciaal
systeem dat ervoor zorgt dat exosomen steeds kleiner worden
gemaakt (en dus geconcentreerder worden). Hierbij hechten clathrin
eiwitten aan het membraan van een exosoom, om er een leeg blaasje
vanaf te halen, om die vervolgens weer naar de trans-kant van het
golgisysteem te brengen.
Synaptic vesicles
Wanneer door de zenuwcel een elektrisch signaal arriveert, versmelten de synaptische blaasjes met
de celmembraan van de zenuwcel, waardoor de neurotransmitters vrijkomen in de synaps. Dit
proces is dus een vorm van exocytose. Na exocytose diffunderen de neurotransmitters door de
synaptische spleet naar receptoren die zich op aangrenzende cellen bevinden, waarna het signaal
verder kan worden verwerkt. Neurotransmitters worden zich vanuit het cytoplasma van de cel naar
de blaasjes getransporteerd door middel van vesiculaire transporteiwitten en actief transport. Dit
systeem werkt erg snel, omdat de
blaasjes worden hergebruikt en dus in
het uiteinde van de synapsen blijven.
Als er steeds nieuwe blaasjes vanuit
het golgi apparaat moesten komen,
zouden elektrische impulsen veel
langzamer overkomen.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
36
Botox en tetanus
Botuline en toxinine zijn proteases die SNARE’s afbreken. Hierdoor verloopt exocytose en endocytose
minder goed. Dit leidt tot het continu samentrekken of verslappen van spieren.
Metabolisme
Stofwisseling, ook wel metabolisme genoemd, is het geheel van biochemische processen die
plaatsvinden in cellen en organismen. Enzymen spelen hierbij een centrale rol.
De stofwisseling heeft onder meer de volgende functies:
De aanmaak van reservestoffen door vastleggen van zonneenergie of chemische energie en het gebruik daarvan
De opname van stoffen
Het vrijmaken van energie uit o.a. opgenomen stoffen
Het gebruik van bouwstoffen en energie als bron voor alle
biologische processen
Het verwerken van afvalstoffen
Een bepaald teveel aan opbouwstoffen elimineren
Hiernaast zie je dat voedselmoleculen kunnen worden omgezet in
nuttige vormen van energie + hitte, en bouwstenen voor
biosynthese (opbouwen van moleculen). Catabolic pathways zijn
afbrekende processen, anabolic pathways zijn de opbouwende
processen.
Lichtreactie
Alle reacties die gebeuren
tijdens de fotosynthese zijn
grofweg te scheiden in twee
reactiewegen. Dit artikel gaat
over de eerste reactieweg, de
lichtreacties. Tijdens de
lichtreacties wordt lichtenergie
omgezet in chemische energie in de vorm van de energiedragende stoffen ATP en NADPH. Hierbij
komt zuurstof vrij als afvalproduct. Tijdens de tweede reactieweg, de calvincyclus (ook wel de
donkerreacties genoemd) worden deze stoffen gebruikt om van koolstofdioxide de energierijke stof
glucose te maken.
De nettoreactie van alle lichtreacties ziet er als volgt uit:
12H2O + 12NADP+ + 18ADP + 18Pi → 6O2 + 12NADPH + 18ATP
Donkerreactie
De reductie door NADPH,H+ gebeurt in meerdere cyclische enzymmatische tussenstappen. Bij
planten vindt de reductie plaats in de stroma van de bladgroenkorrels. De ontdekkers zijn de NoordAmerikaanse chemicus Melvin Calvin en Andy Benson van de Universiteit van Berkeley in Californië,
waaraan de reactie de naam te danken heeft.
Nettoreactie Calvincyclus:
6CO2+ 12 NADPH,H++ 18ATP →C6H12O6+ 6H2O + 12 NADP+ + 18 ADP + 18 Pi
Dit resulteert in de volgende reactie: 12H2O + 6CO2 + licht → C6H12O6 (glucose) + 6O2 + 6H2O
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
37
Fotosynthese en dissimilatie
Je zou grofweg kunnen zeggen het
doel van de fotosynthese, het
vastleggen van energie in glucose is.
Eukaryoten breken suikers af, om de
energie die daarbij vrijkomt te
kunnen gebruiken. Hierbij is zuurstof
nodig.
Koolstofcyclus
De koolstofkringloop is de bekendste biogeochemische kringloop en beschrijft alle processen
waarmee het element koolstof door het systeem Aarde circuleert. De koolstofkringloop beschrijft
onder andere wat er gebeurt met door mensen uitgestote koolstofdioxide in de atmosfeer. Omdat
koolstofdioxide een belangrijk broeikasgas is, vormen de onduidelijkheden binnen de
koolstofkringloop een belangrijk onderzoeksterrein binnen de biochemie, geochemie en
klimatologie.
NADPH
Het komt voor in 2 soorten: NAD+ en NADP+ dat een
extra veresterde fosfaatgroep op de 2'-OH-groep van
de adenosine draagt. Het verschil tussen beide
moleculen ligt bij de functie ervan. Meestal fungeert
NAD+ als cofactor bij katabole reacties (afbraak),
terwijl NADP+ dienst doet bij anabole reacties
(opbouw). Daarnaast wordt NAD+ bij dissimilatie
gebruikt, en NADP+ bij fotosynthese.
NADH is de gereduceerde vorm van NAD+, en NADPH
is de gereduceerde vorm van NADP+. De met een
proton en elektron opgeladen moleculenzijn dan
NADH en NADPH die optreden als elektronendonor.
Deze elektronendonoren dienen als een soort van
mobiele energiestockage. Die energie komt terug
vrij wanneer NADH of NADPH zich terug omgezet
wordt naar NAD+ en NADP+. In dit tentamen zullen
we vooral letten op NADP+ en NADPH+H+.
Wanneer NADPH zijn energie kwijt wil zal de
volgende reactie lopen:
NADPH NADP+ + H+ + 2e-
Energie leverende voedingsstoffen
- Suikers: afbraak d.m.v. glycolyse tot pyruvaat in het cytosol. Pyruvaat wordt in mitochondrion
omgezet tot acetyl CoA
- Acetyl CoA wordt afgebroken via de citroenzuurcyclus in de matrix van het mitochondrion. Hierbij
wordt ATP gesynthetiseerd
- Vetten: afbraak tot vetzuren en vervolgens tot acetyl CoA.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
38
Citroenzuurcyclus
Het doel van de citroenzuurcyclus is het vormen van
NADH en CO2 uit acetyl CoA. Wanneer de
citroenzuurcyclus één keer is doorlopen, zullen er 3
NADH-moleculen, één GTP molecuul en twee CO2
moleculen gevormd zijn. Zoals hierboven beschreven
staat wordt suiker eerst afgebroken tot acetyl CoA,
daarbij worden veel bruikbare tussenproducten gevormd,
die weer een ander doel hebben in een ander
systeem/complex.
Oxidatieve fosforylering/ATP synthase
Zoals ik al zei worden tijdens de citroenzuurcyclus (die zich in de matrix van het mitochondrion
afspeelt) NADH en elektronen gevormd. Deze producten van de citroenzuurcyclus gaan we nu
gebruiken om ATP te vormen. Volgens de chemiosmotische theorie werkt de oxfos (oxidatieve
fosforylering) volgens de volgende principes.
energierijke elektronen uit de citroenzuurcyclus geven hun energie af in een serie
redoxreacties, waarbij uiteindelijk zuurstof wordt gereduceerd tot water.
Deze energie wordt gebruikt om een protonengradiënt te creëren.
Deze protonengradiënt drijft de
synthese van ATP aan.
Het de twee elektronen die vrijkomen tijdens
de citroenzuurcyclus hebben zoveel energie,
dat ze 3 fotosystemen aan kunnen zetten tot
het transporteren van H+-ionen van het
matrix, naar het tussenmembraan. Hierdoor
ontstaat zowel een protonen gradiënt als een
lading gradiënt. Door dit verschil, zal er altijd
H+ naar de matrix van het mitochondrion
willen stromen. Hier komt ATP synthase goed
van pas. Door het verschil in concentratie H+,
zal het gamma eiwit in ATP synthase gaan
draaien als een soort rotor. Hierdoor zullen
ADP en P samen worden geperst tot ATP. Je
kunt dus zeggen dat de mechanische kracht
ATP vormt (bestudeer ook de afbeelding voor
het tentamen).
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
39
Cristae
De binnenste mitochondriale membraan is verdeeld in
talrijke plooien of invaginaties die we cristae noemen.
Deze vergroten de oppervlakte van de binnenste
mitochondriale membraan en daarmee het vermogen om
ATP te produceren. Voor typische levermitochondriën is
het gebied van de binnenste membraan ongeveer vijf keer
groter dan de buitenste membraan. Deze verhouding is
variabel en de mitochondriën van de cellen die een
grotere vraag naar ATP hebben, zoals spiercellen,
bevatten nog meer cristae. In de afbeelding hiernaast zijn
de ATP synthase moleculen met pijltjes aangegeven.
Fotosynthese
In een chloroplast verlopen bijna dezelfde processen. Het
verschil hierin is goed op de verschillende DIA’s in de
map te zien. Je hoeft deze niet door en door te
kennen, maar het is wel belangrijk dat je de
verschillen kan herkennen. Ook is het belangrijk dat
je weet hoe een chloroplast en een mitochondrion er
van buiten en van binnen uitzien. Ook moet je weten
waar in het organel (zowel in een chloroplast als een
mitochondrion) de pH hoger is, en waar het
membraan positief of negatief geladen is. Ook moet
je weten waar elk proces zich afspeelt, (bijvoorbeeld
de koolstoffixatie reacties in het stroma).
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
40
Mitochondriën en Chloroplasten (kennen voor tentamen)
Overeenkomsten:
- Vorming proton gradiënt d.m.v. elektronen transportketen.
- ATP synthese door terugstroom protonen
door ATP-synthase .
- Dubbele membraan
- Eigen genoom
- Ribosomen
- Energie conversie
- Elektronen transport keten
-ATP synthase
Verschillen:
- Mitochondriën halen hun energie uit
verbranding en chloroplasten uit zonlicht.
- Mitochondriën consumeren O2 en
chloroplasten produceren O2.
- Mitochondriën produceren CO2 en chloroplasten consumeren CO2.
Samenvatting Celbiologie (2011-2012)
Rijksuniversiteit Groningen
Niek Groot
41
lipiden")